

Redox Sensing by Fe2+ in Bacterial Fur Family Metalloregulators
- PMID: 28938859
- PMCID: PMC6217742
- DOI: 10.1089/ars.2017.7359
Redox Sensing by Fe2+ in Bacterial Fur Family Metalloregulators
Abstract
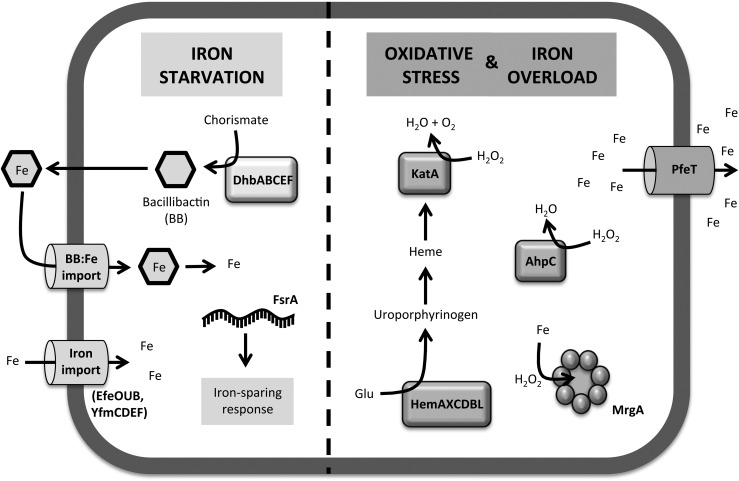
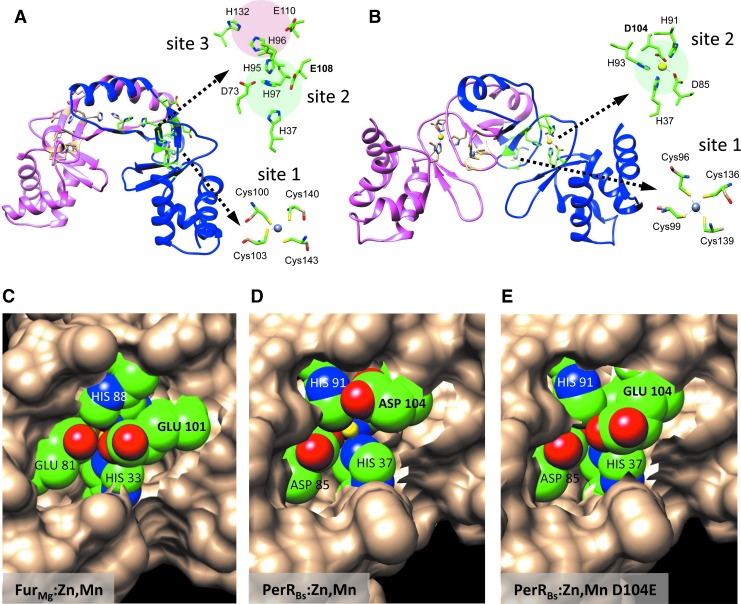
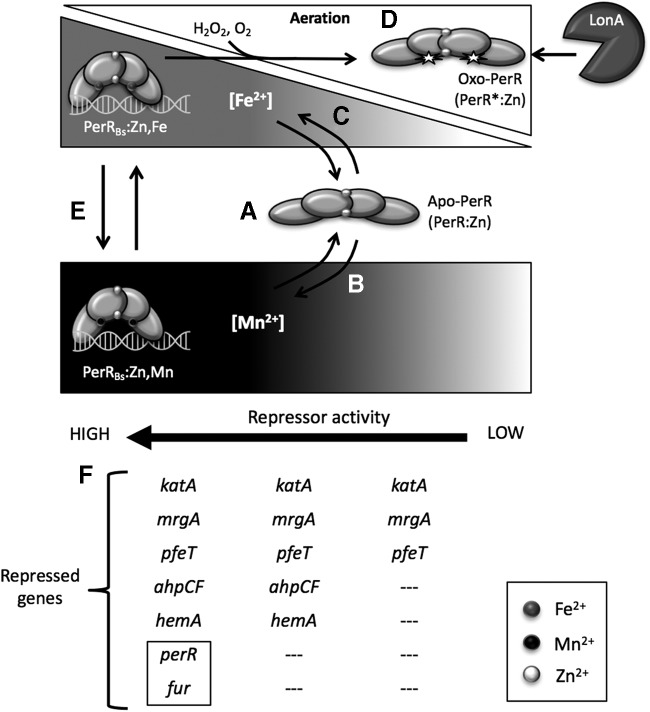
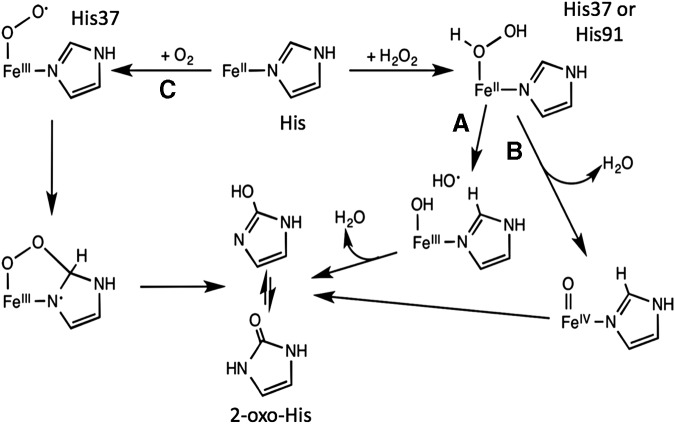
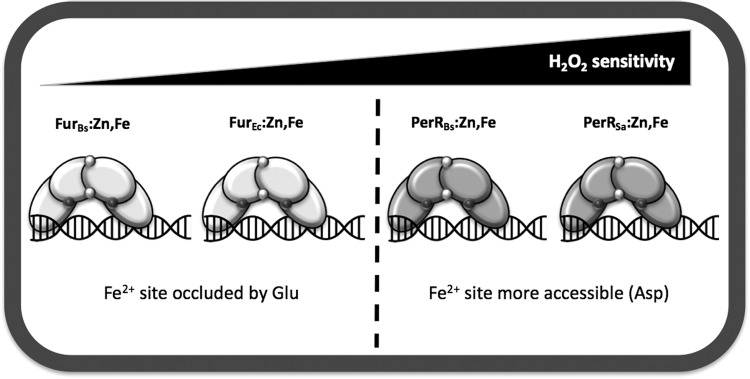
Significance: Iron is required for growth and is often redox active under cytosolic conditions. As a result of its facile redox chemistry, iron homeostasis is intricately involved with oxidative stress. Bacterial adaptation to iron limitation and oxidative stress often involves ferric uptake regulator (Fur) proteins: a diverse set of divalent cation-dependent, DNA-binding proteins that vary widely in both metal selectivity and sensitivity to metal-catalyzed oxidation. Recent Advances: Bacteria contain two Fur family metalloregulators that use ferrous iron (Fe2+) as their cofactor, Fur and PerR. Fur functions to regulate iron homeostasis in response to changes in intracellular levels of Fe2+. PerR also binds Fe2+, which enables metal-catalyzed protein oxidation as a mechanism for sensing hydrogen peroxide (H2O2).
Critical issues: To effectively regulate iron homeostasis, Fur has an Fe2+ affinity tuned to monitor the labile iron pool of the cell and may be under selective pressure to minimize iron oxidation, which would otherwise lead to an inappropriate increase in iron uptake under oxidative stress conditions. Conversely, Fe2+ is bound more tightly to PerR but exhibits high H2O2 reactivity, which enables a rapid induction of peroxide stress genes.
Future directions: The features that determine the disparate reactivity of these proteins with oxidants are still poorly understood. A controlled, comparative analysis of the affinities of Fur/PerR proteins for their metal cofactors and their rate of reactivity with H2O2, combined with structure/function analyses, will be needed to define the molecular mechanisms that have facilitated this divergence of function between these two paralogous regulators.
Keywords: Bacillus subtilis; Staphylococcus aureus; hydrogen peroxide; iron; metalloregulation; oxidative stress.
Figures
References
-
- Agriesti F, Roncarati D, Musiani F, Del Campo C, Iurlaro M, Sparla F, Ciurli S, Danielli A, and Scarlato V. FeON-FeOFF: the Helicobacter pylori Fur regulator commutates iron-responsive transcription by discriminative readout of opposed DNA grooves. Nucleic Acids Res 42: 3138–3151, 2014 - PMC - PubMed
-
- Ahn BE, Cha J, Lee EJ, Han AR, Thompson CJ, and Roe JH. Nur, a nickel-responsive regulator of the Fur family, regulates superoxide dismutases and nickel transport in Streptomyces coelicolor. Mol Microbiol 59: 1848–1858, 2006 - PubMed
-
- Althaus EW, Outten CE, Olson KE, Cao H, and O'Halloran TV. The ferric uptake regulation (Fur) repressor is a zinc metalloprotein. Biochemistry 38: 6559–6569, 1999 - PubMed
-
- Anbar AD. Elements and evolution. Science 322: 1481–1483, 2008 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
