

Different developmental histories of beta-cells generate functional and proliferative heterogeneity during islet growth
- PMID: 28939870
- PMCID: PMC5610262
- DOI: 10.1038/s41467-017-00461-3
Different developmental histories of beta-cells generate functional and proliferative heterogeneity during islet growth
Abstract
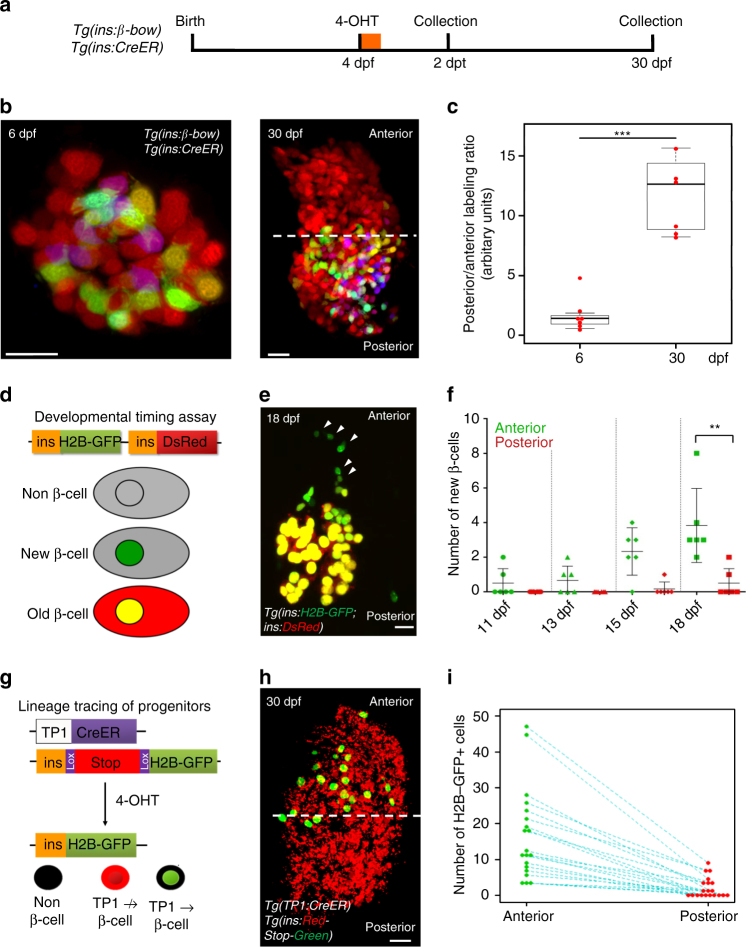
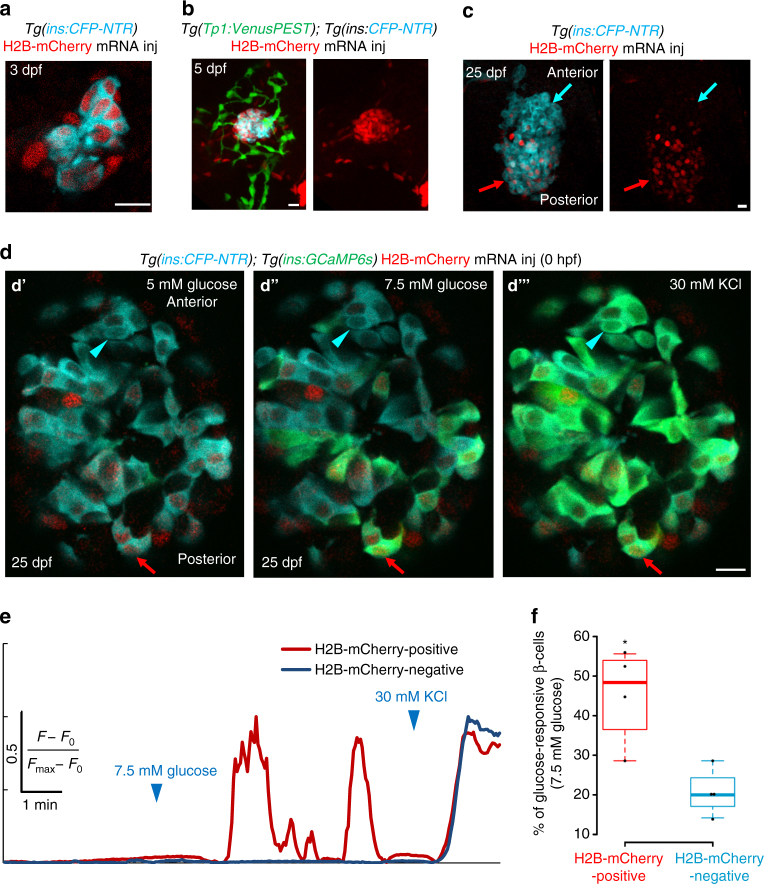
The proliferative and functional heterogeneity among seemingly uniform cells is a universal phenomenon. Identifying the underlying factors requires single-cell analysis of function and proliferation. Here we show that the pancreatic beta-cells in zebrafish exhibit different growth-promoting and functional properties, which in part reflect differences in the time elapsed since birth of the cells. Calcium imaging shows that the beta-cells in the embryonic islet become functional during early zebrafish development. At later stages, younger beta-cells join the islet following differentiation from post-embryonic progenitors. Notably, the older and younger beta-cells occupy different regions within the islet, which generates topological asymmetries in glucose responsiveness and proliferation. Specifically, the older beta-cells exhibit robust glucose responsiveness, whereas younger beta-cells are more proliferative but less functional. As the islet approaches its mature state, heterogeneity diminishes and beta-cells synchronize function and proliferation. Our work illustrates a dynamic model of heterogeneity based on evolving proliferative and functional beta-cell states.Βeta-cells have recently been shown to be heterogeneous with regard to morphology and function. Here, the authors show that β-cells in zebrafish switch from proliferative to functional states with increasing time since β-cell birth, leading to functional and proliferative heterogeneity.
Conflict of interest statement
The authors declare no competing financial interests.
Figures





Similar articles
-
In vivo imaging of β-cell function reveals glucose-mediated heterogeneity of β-cell functional development.Elife. 2019 Jan 29;8:e41540. doi: 10.7554/eLife.41540. Elife. 2019. PMID: 30694176 Free PMC article.
-
Single-cell analyses identify distinct and intermediate states of zebrafish pancreatic islet development.J Mol Cell Biol. 2019 Jun 1;11(6):435-447. doi: 10.1093/jmcb/mjy064. J Mol Cell Biol. 2019. PMID: 30407522 Free PMC article.
-
An insulin signaling feedback loop regulates pancreas progenitor cell differentiation during islet development and regeneration.Dev Biol. 2016 Jan 15;409(2):354-69. doi: 10.1016/j.ydbio.2015.12.003. Epub 2015 Dec 3. Dev Biol. 2016. PMID: 26658317 Free PMC article.
-
Is a β cell a β cell?Curr Opin Endocrinol Diabetes Obes. 2017 Apr;24(2):92-97. doi: 10.1097/MED.0000000000000322. Curr Opin Endocrinol Diabetes Obes. 2017. PMID: 28099206 Free PMC article. Review.
-
Impact of islet architecture on β-cell heterogeneity, plasticity and function.Nat Rev Endocrinol. 2016 Dec;12(12):695-709. doi: 10.1038/nrendo.2016.147. Epub 2016 Sep 2. Nat Rev Endocrinol. 2016. PMID: 27585958 Review.
Cited by
-
In vivo imaging of β-cell function reveals glucose-mediated heterogeneity of β-cell functional development.Elife. 2019 Jan 29;8:e41540. doi: 10.7554/eLife.41540. Elife. 2019. PMID: 30694176 Free PMC article.
-
Beta-cell excitability and excitability-driven diabetes in adult Zebrafish islets.Physiol Rep. 2019 Jun;7(11):e14101. doi: 10.14814/phy2.14101. Physiol Rep. 2019. PMID: 31161721 Free PMC article.
-
Developmental beta-cell death orchestrates the islet's inflammatory milieu by regulating immune system crosstalk.EMBO J. 2025 Feb;44(4):1131-1153. doi: 10.1038/s44318-024-00332-w. Epub 2025 Jan 6. EMBO J. 2025. PMID: 39762647 Free PMC article.
-
The role of beta cell heterogeneity in islet function and insulin release.J Mol Endocrinol. 2018 Jul;61(1):R43-R60. doi: 10.1530/JME-18-0011. Epub 2018 Apr 16. J Mol Endocrinol. 2018. PMID: 29661799 Free PMC article. Review.
-
Pancreatic exocrine damage induces beta cell stress in zebrafish larvae.Diabetologia. 2025 Aug;68(8):1754-1768. doi: 10.1007/s00125-025-06432-4. Epub 2025 Apr 28. Diabetologia. 2025. PMID: 40295334 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
