

A Single Sphingomyelin Species Promotes Exosomal Release of Endoglin into the Maternal Circulation in Preeclampsia
- PMID: 28939895
- PMCID: PMC5610344
- DOI: 10.1038/s41598-017-12491-4
A Single Sphingomyelin Species Promotes Exosomal Release of Endoglin into the Maternal Circulation in Preeclampsia
Abstract
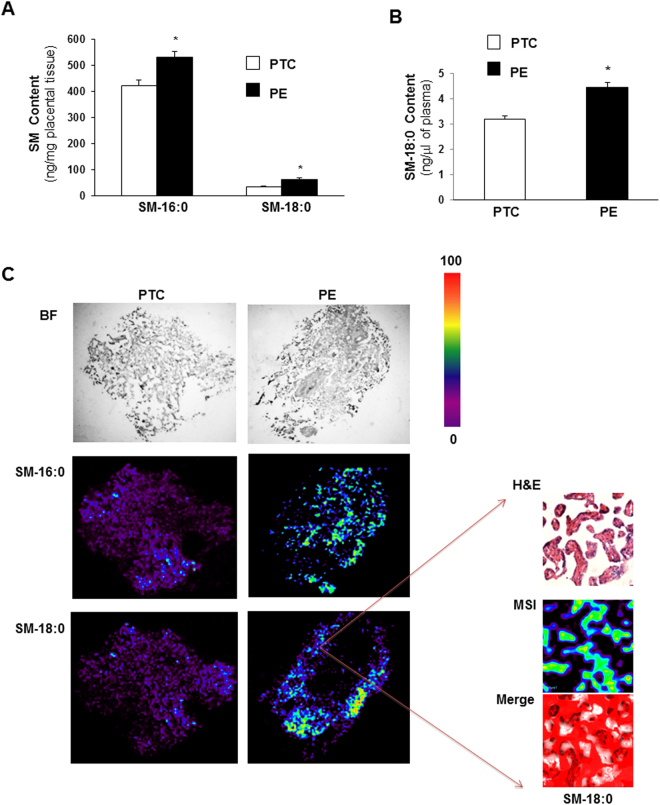
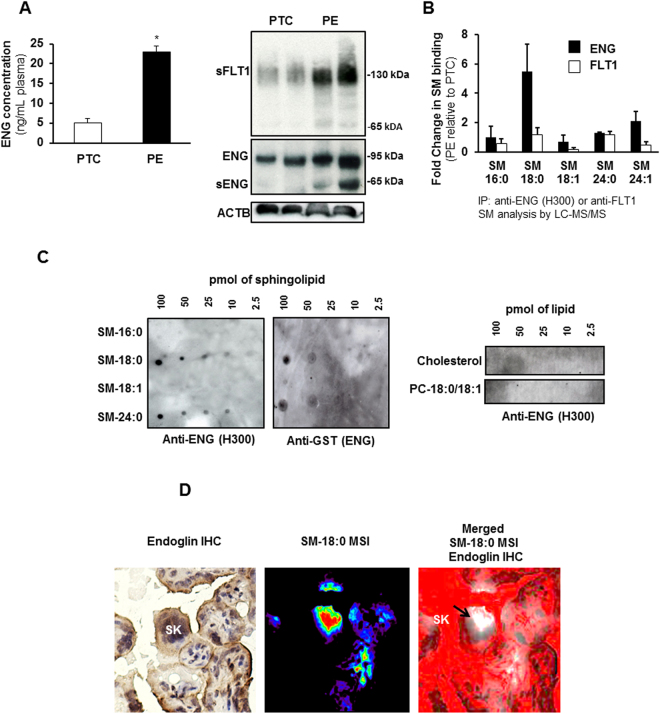
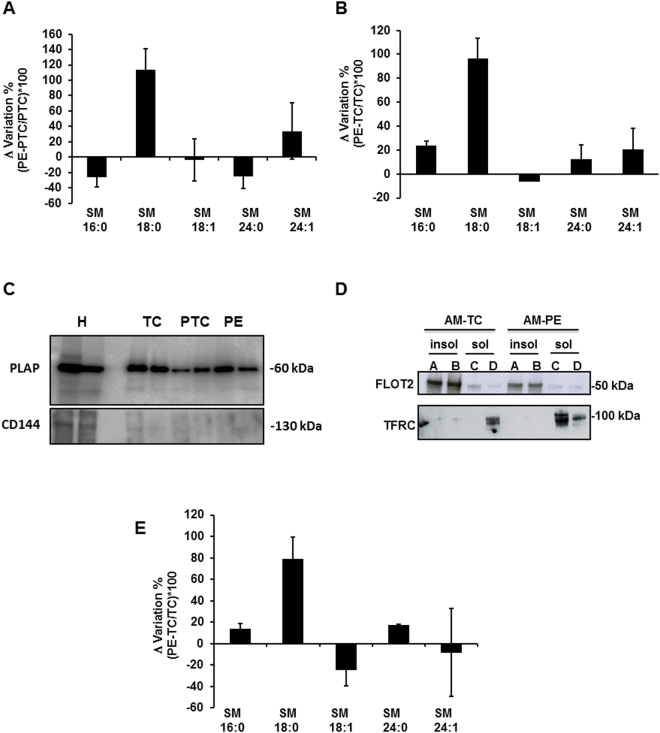
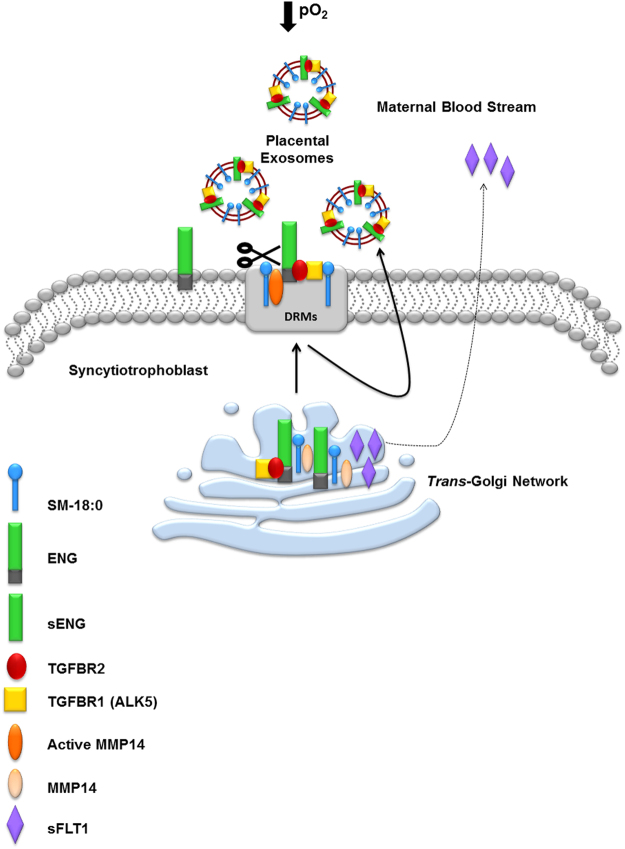
Preeclampsia (PE), an hypertensive disorder of pregnancy, exhibits increased circulating levels of a short form of the auxillary TGF-beta (TGFB) receptor endoglin (sENG). Until now, its release and functionality in PE remains poorly understood. Here we show that ENG selectively interacts with sphingomyelin(SM)-18:0 which promotes its clustering with metalloproteinase 14 (MMP14) in SM-18:0 enriched lipid rafts of the apical syncytial membranes from PE placenta where ENG is cleaved by MMP14 into sENG. The SM-18:0 enriched lipid rafts also contain type 1 and 2 TGFB receptors (TGFBR1 and TGFBR2), but not soluble fms-like tyrosine kinase 1 (sFLT1), another protein secreted in excess in the circulation of women with PE. The truncated ENG is then released into the maternal circulation via SM-18:0 enriched exosomes together with TGFBR1 and 2. Such an exosomal TGFB receptor complex could be functionally active and block the vascular effects of TGFB in the circulation of PE women.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
