

Ghrelin receptor antagonism of hyperlocomotion in cocaine-sensitized mice requires βarrestin-2
- PMID: 28941296
- PMCID: PMC5705267
- DOI: 10.1002/syn.22012
Ghrelin receptor antagonism of hyperlocomotion in cocaine-sensitized mice requires βarrestin-2
Abstract
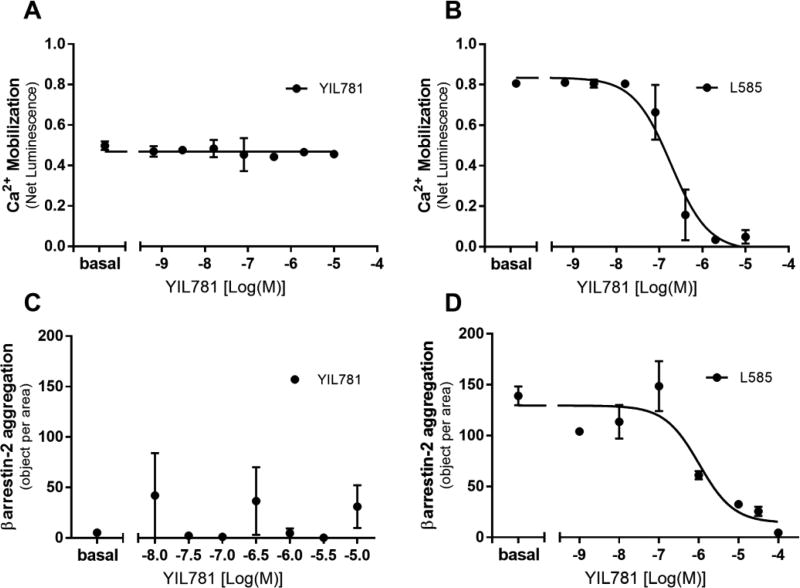
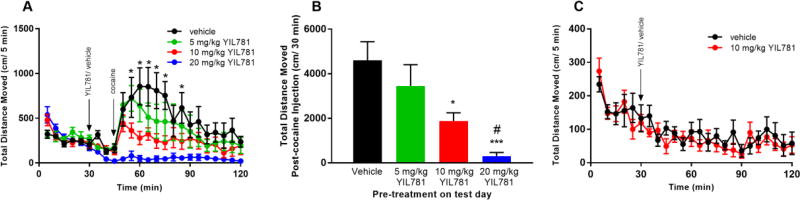
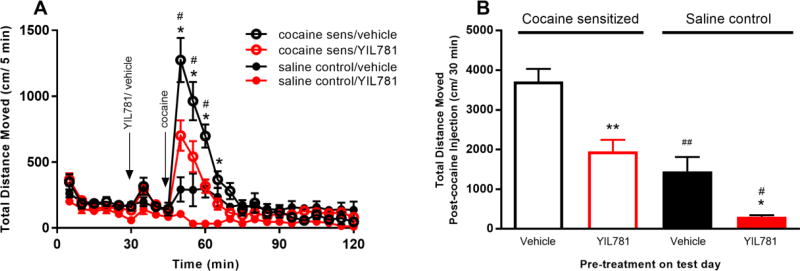
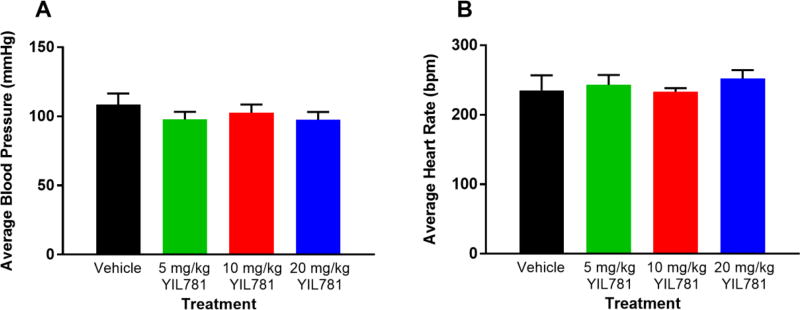
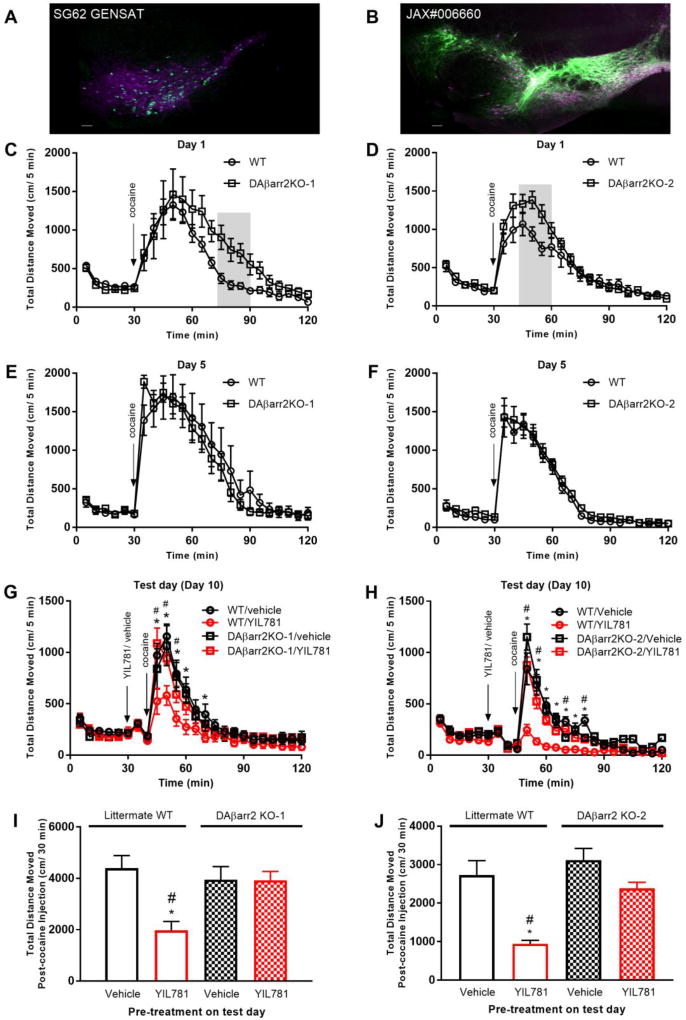
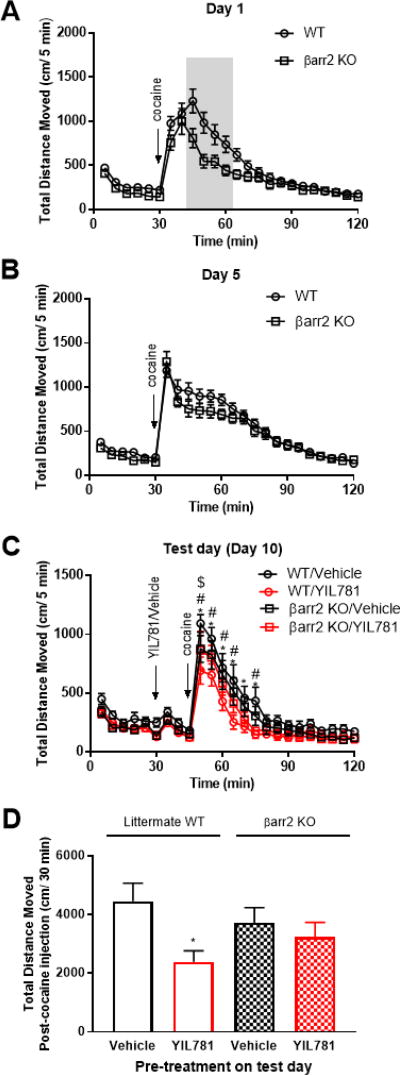
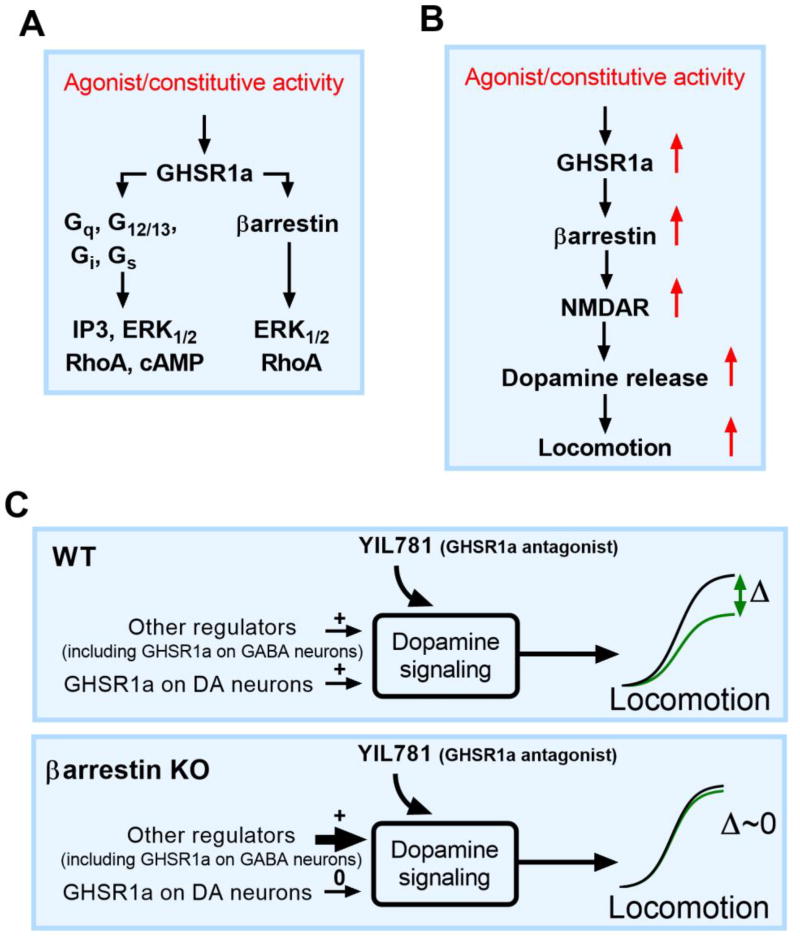
The "brain-gut" peptide ghrelin, which mediates food-seeking behaviors, is recognized as a very strong endogenous modulator of dopamine (DA) signaling. Ghrelin binds the G protein-coupled receptor GHSR1a, and administration of ghrelin increases the rewarding properties of psychostimulants while ghrelin receptor antagonists decrease them. In addition, the GHSR1a signals through βarrestin-2 to regulate actin/stress fiber rearrangement, suggesting βarrestin-2 participation in the regulation of actin-mediated synaptic plasticity for addictive substances like cocaine. The effects of ghrelin receptor ligands on reward strongly suggest that modulation of ghrelin signaling could provide an effective strategy to ameliorate undesirable behaviors arising from addiction. To investigate this possibility, we tested the effects of ghrelin receptor antagonism in a cocaine behavioral sensitization paradigm using DA neuron-specific βarrestin-2 KO mice. Our results show that these mice sensitize to cocaine as well as wild-type littermates. The βarrestin-2 KO mice, however, no longer respond to the locomotor attenuating effects of the GHSR1a antagonist YIL781. The data presented here suggest that the separate stages of addictive behavior differ in their requirements for βarrestin-2 and show that pharmacological inhibition of βarrestin-2 function through GHSR1a antagonism is not equivalent to the loss of βarrestin-2 function achieved by genetic ablation. These data support targeting GHSR1a signaling in addiction therapy but indicate that using signaling biased compounds that modulate βarrestin-2 activity differentially from G protein activity may be required.
Keywords: YIL781; addiction; arrestin; cocaine; ghrelin.
© 2017 Wiley Periodicals, Inc.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Black JW, Leff P. Operational models of pharmacological agonism. Proc R Soc Lond B Biol Sci. 1983;220(1219):141–162. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials