

Transcriptional Architecture of Synaptic Communication Delineates GABAergic Neuron Identity
- PMID: 28942923
- PMCID: PMC5772785
- DOI: 10.1016/j.cell.2017.08.032
Transcriptional Architecture of Synaptic Communication Delineates GABAergic Neuron Identity
Abstract
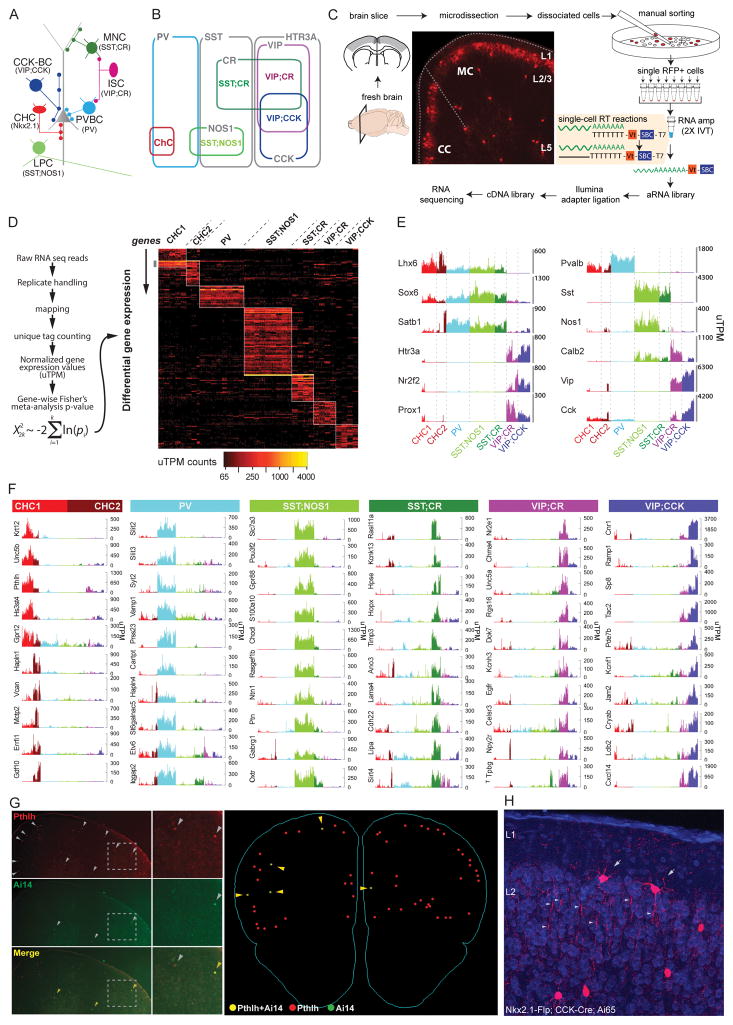
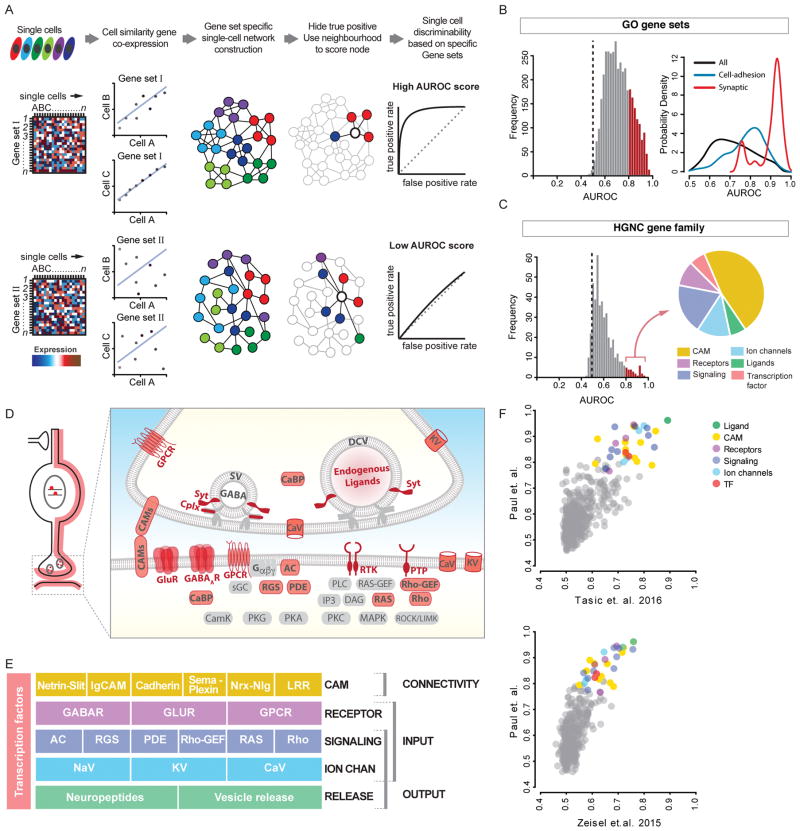
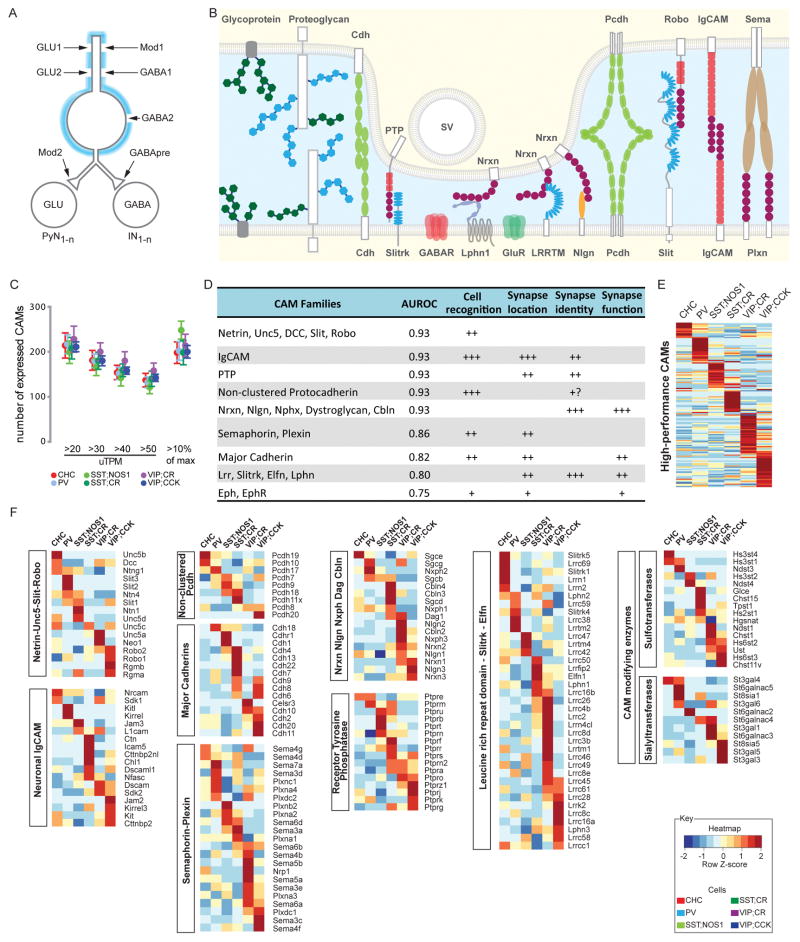
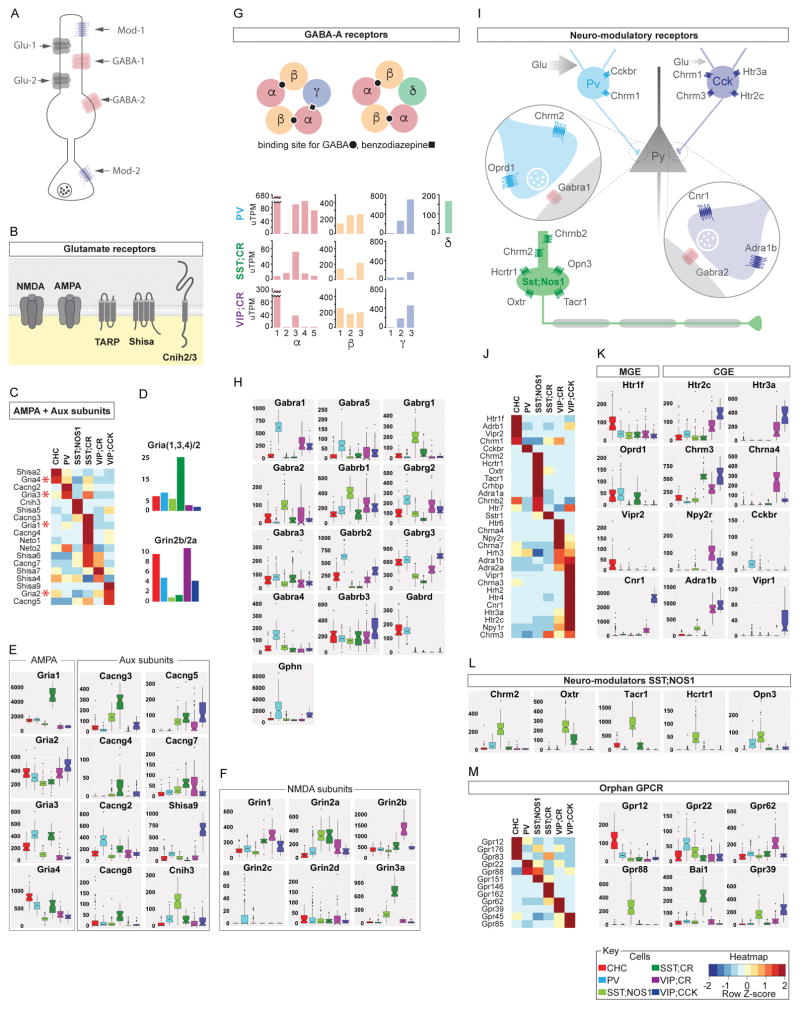
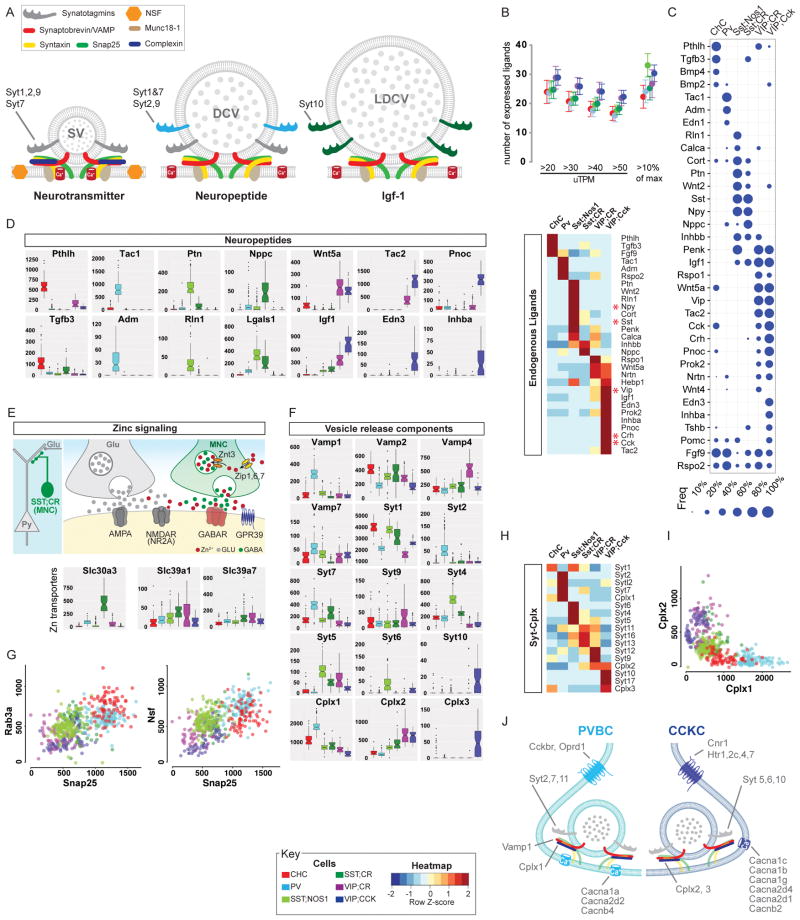
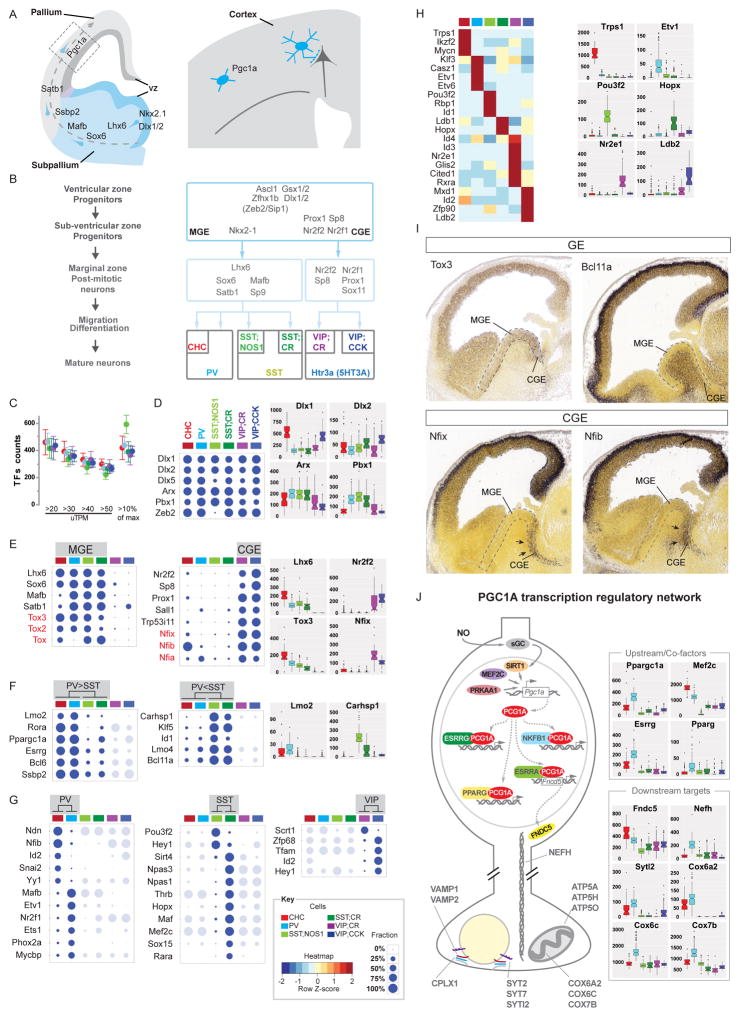
Understanding the organizational logic of neural circuits requires deciphering the biological basis of neuronal diversity and identity, but there is no consensus on how neuron types should be defined. We analyzed single-cell transcriptomes of a set of anatomically and physiologically characterized cortical GABAergic neurons and conducted a computational genomic screen for transcriptional profiles that distinguish them from one another. We discovered that cardinal GABAergic neuron types are delineated by a transcriptional architecture that encodes their synaptic communication patterns. This architecture comprises 6 categories of ∼40 gene families, including cell-adhesion molecules, transmitter-modulator receptors, ion channels, signaling proteins, neuropeptides and vesicular release components, and transcription factors. Combinatorial expression of select members across families shapes a multi-layered molecular scaffold along the cell membrane that may customize synaptic connectivity patterns and input-output signaling properties. This molecular genetic framework of neuronal identity integrates cell phenotypes along multiple axes and provides a foundation for discovering and classifying neuron types.
Keywords: GABAergic interneurons; MetaNeighbor; cell type; co-expression; gene family; network analysis; neuronal identity; single-cell transcriptomes; synaptic communication; transcriptional architecture.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures

References
-
- Alberts B, Johnson A, Lewis J, Morgan D, Raff M, Roberts K, Walter P. Cell Signaling Molecular Biology of the Cell. 2014;Chapter 15
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
