

EHD Proteins Cooperate to Generate Caveolar Clusters and to Maintain Caveolae during Repeated Mechanical Stress
- PMID: 28943089
- PMCID: PMC5640515
- DOI: 10.1016/j.cub.2017.07.047
EHD Proteins Cooperate to Generate Caveolar Clusters and to Maintain Caveolae during Repeated Mechanical Stress
Abstract
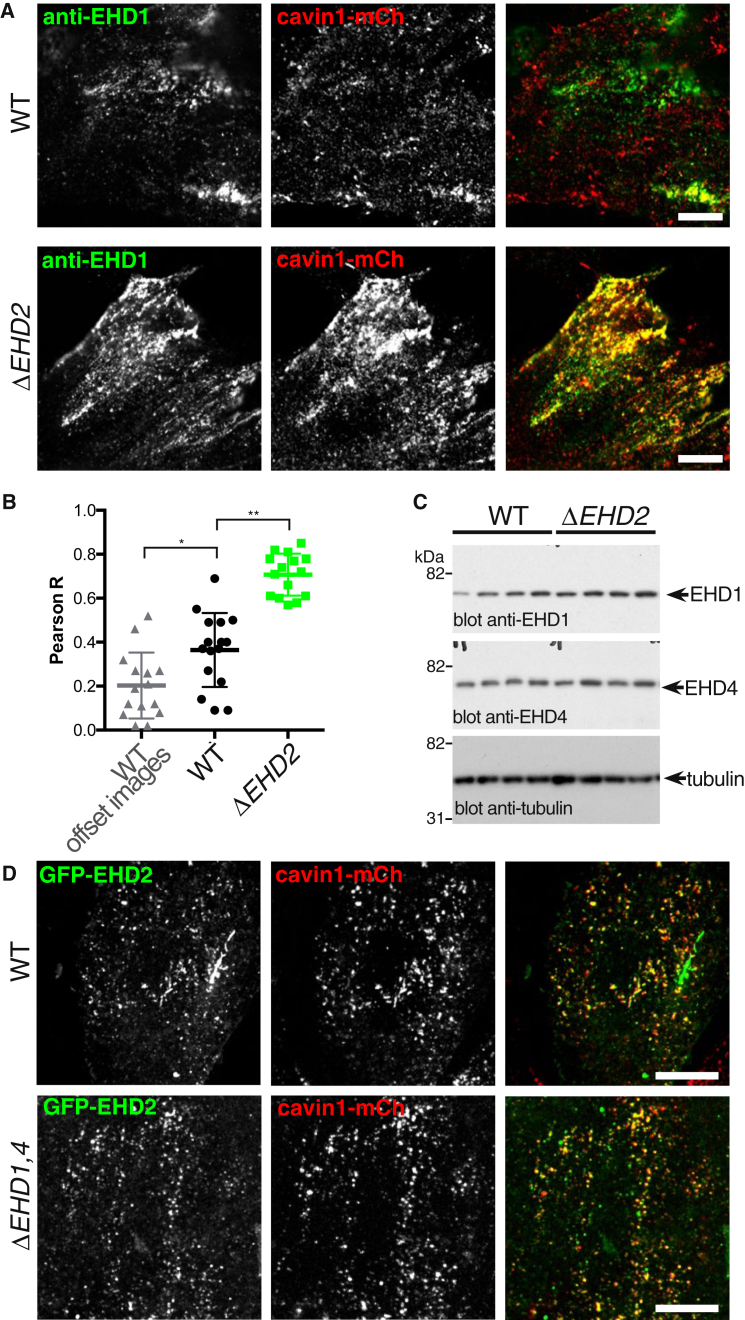
Caveolae introduce flask-shaped convolutions into the plasma membrane and help to protect the plasma membrane from damage under stretch forces. The protein components that form the bulb of caveolae are increasingly well characterized, but less is known about the contribution of proteins that localize to the constricted neck. Here we make extensive use of multiple CRISPR/Cas9-generated gene knockout and knockin cell lines to investigate the role of Eps15 Homology Domain (EHD) proteins at the neck of caveolae. We show that EHD1, EHD2, and EHD4 are recruited to caveolae. Recruitment of the other EHDs increases markedly when EHD2, which has been previously detected at caveolae, is absent. Construction of knockout cell lines lacking EHDs 1, 2, and 4 confirms this apparent functional redundancy. Two striking sets of phenotypes are observed in EHD1,2,4 knockout cells: (1) the characteristic clustering of caveolae into higher-order assemblies is absent; and (2) when the EHD1,2,4 knockout cells are subjected to prolonged cycles of stretch forces, caveolae are destabilized and the plasma membrane is prone to rupture. Our data identify the first molecular components that act to cluster caveolae into a membrane ultrastructure with the potential to extend stretch-buffering capacity and support a revised model for the function of EHDs at the caveolar neck.
Keywords: EHD2; caveolae; caveolin; cavin; cell; membrane; stretch.
Copyright © 2017 MRC Laboratory of Molecular Biology. Published by Elsevier Ltd.. All rights reserved.
Figures






Similar articles
-
Eps15 Homology Domain Protein 4 (EHD4) is required for Eps15 Homology Domain Protein 1 (EHD1)-mediated endosomal recruitment and fission.PLoS One. 2020 Sep 23;15(9):e0239657. doi: 10.1371/journal.pone.0239657. eCollection 2020. PLoS One. 2020. PMID: 32966336 Free PMC article.
-
Differential roles of C-terminal Eps15 homology domain proteins as vesiculators and tubulators of recycling endosomes.J Biol Chem. 2013 Oct 18;288(42):30172-30180. doi: 10.1074/jbc.M113.488627. Epub 2013 Sep 9. J Biol Chem. 2013. PMID: 24019528 Free PMC article.
-
EHD2 regulates caveolar dynamics via ATP-driven targeting and oligomerization.Mol Biol Cell. 2012 Apr;23(7):1316-29. doi: 10.1091/mbc.E11-09-0787. Epub 2012 Feb 9. Mol Biol Cell. 2012. PMID: 22323287 Free PMC article.
-
The role of membrane-shaping BAR domain proteins in caveolar invagination: from mechanistic insights to pathophysiological consequences.Biochem Soc Trans. 2020 Feb 28;48(1):137-146. doi: 10.1042/BST20190377. Biochem Soc Trans. 2020. PMID: 32104881 Review.
-
Scratching the surface: actin' and other roles for the C-terminal Eps15 homology domain protein, EHD2.Histol Histopathol. 2014 Mar;29(3):285-92. doi: 10.14670/HH-29.285. Epub 2013 Dec 18. Histol Histopathol. 2014. PMID: 24347515 Free PMC article. Review.
Cited by
-
Lung Endothelial Transcytosis.Compr Physiol. 2020 Mar 12;10(2):491-508. doi: 10.1002/cphy.c190012. Compr Physiol. 2020. PMID: 32163197 Free PMC article. Review.
-
EHD2-mediated restriction of caveolar dynamics regulates cellular fatty acid uptake.Proc Natl Acad Sci U S A. 2020 Mar 31;117(13):7471-7481. doi: 10.1073/pnas.1918415117. Epub 2020 Mar 13. Proc Natl Acad Sci U S A. 2020. PMID: 32170013 Free PMC article.
-
EHD2 is a Predictive Biomarker of Chemotherapy Efficacy in Triple Negative Breast Carcinoma.Sci Rep. 2020 May 14;10(1):7998. doi: 10.1038/s41598-020-65054-5. Sci Rep. 2020. PMID: 32409676 Free PMC article.
-
ATP-dependent membrane remodeling links EHD1 functions to endocytic recycling.Nat Commun. 2018 Dec 5;9(1):5187. doi: 10.1038/s41467-018-07586-z. Nat Commun. 2018. PMID: 30518883 Free PMC article.
-
Caveolin-1 Regulates Atherogenesis by Attenuating Low-Density Lipoprotein Transcytosis and Vascular Inflammation Independently of Endothelial Nitric Oxide Synthase Activation.Circulation. 2019 Jul 16;140(3):225-239. doi: 10.1161/CIRCULATIONAHA.118.038571. Epub 2019 Jun 3. Circulation. 2019. PMID: 31154825 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
