

Active Mechanisms of Vibration Encoding and Frequency Filtering in Central Mechanosensory Neurons
- PMID: 28943231
- PMCID: PMC5771506
- DOI: 10.1016/j.neuron.2017.09.004
Active Mechanisms of Vibration Encoding and Frequency Filtering in Central Mechanosensory Neurons
Abstract
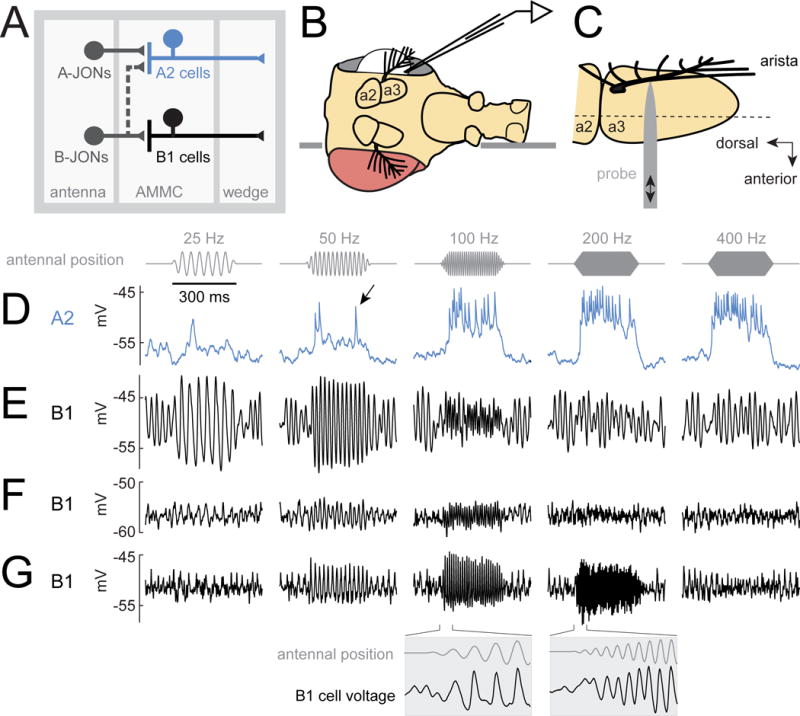
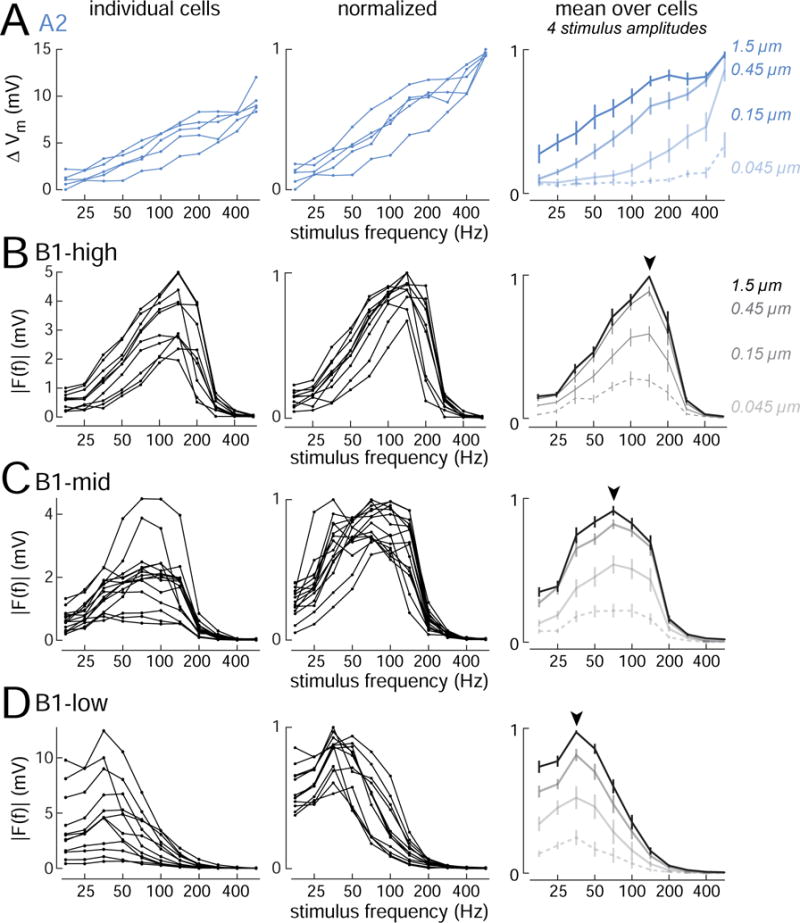
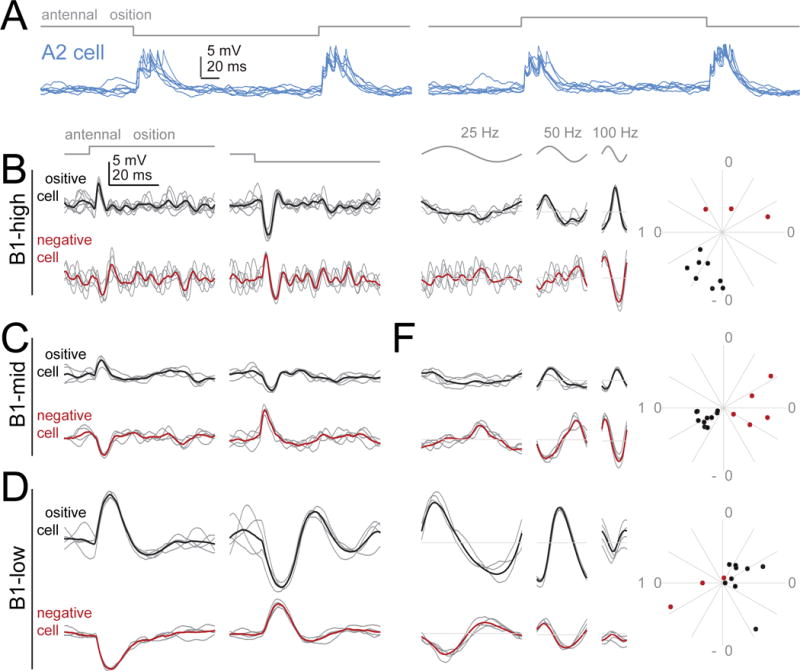
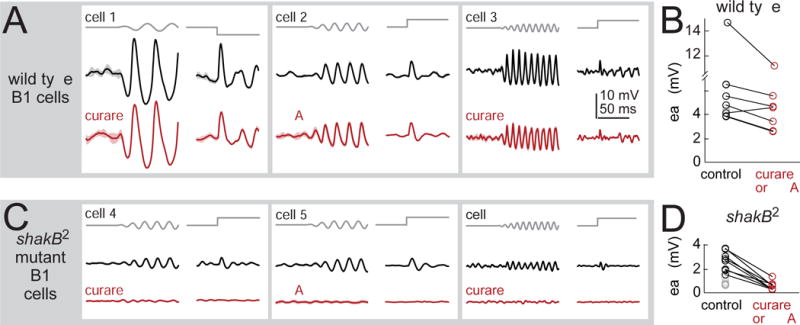
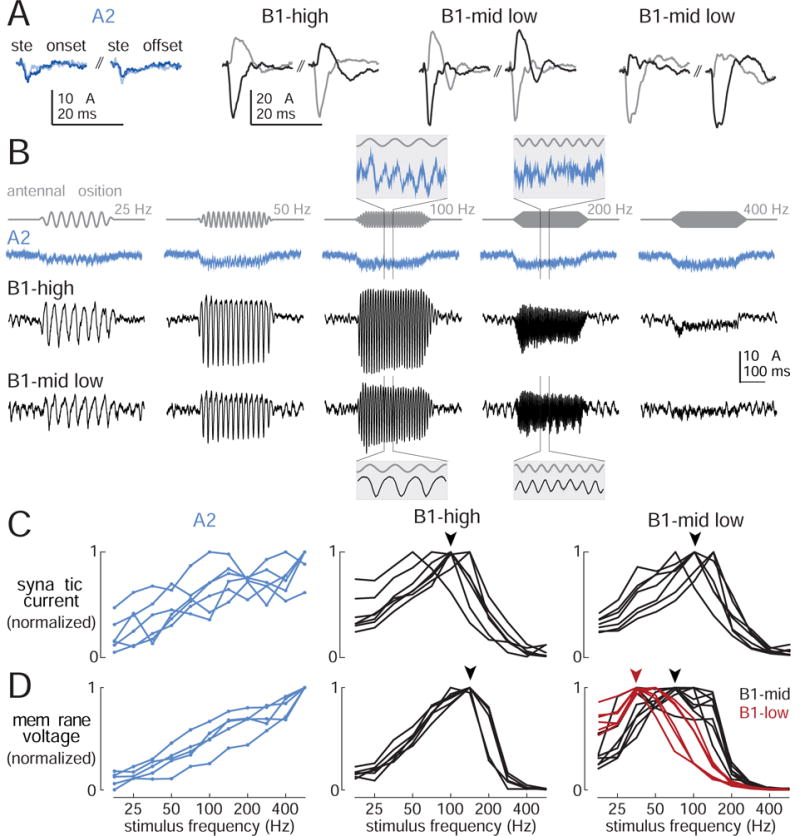
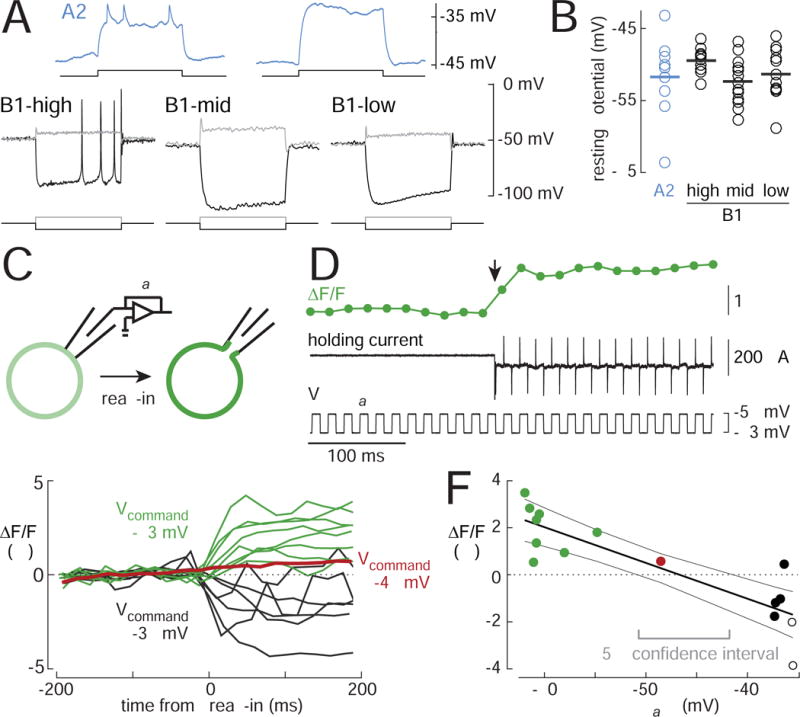
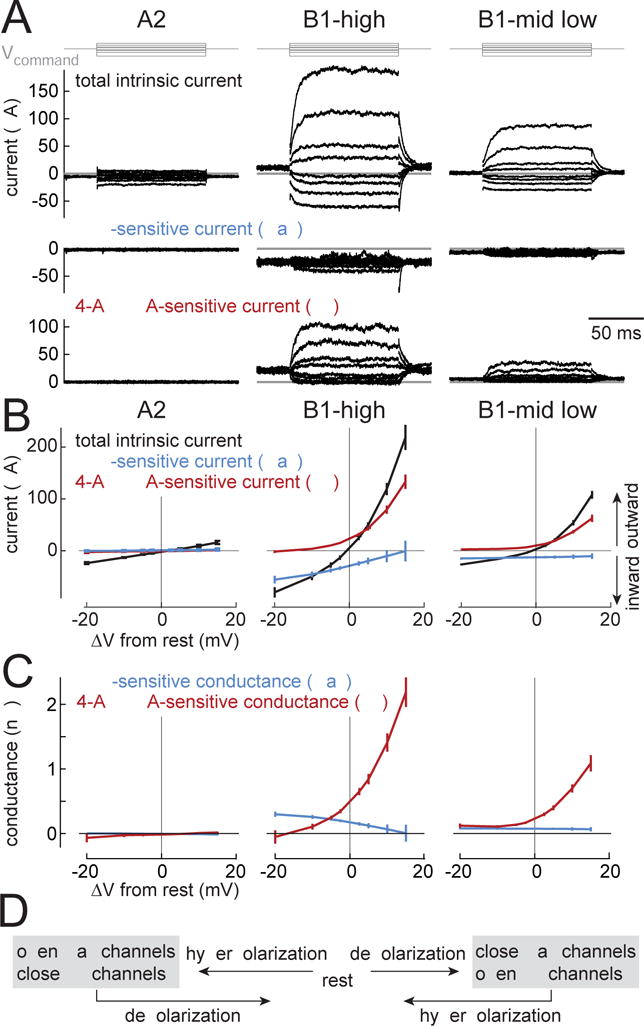
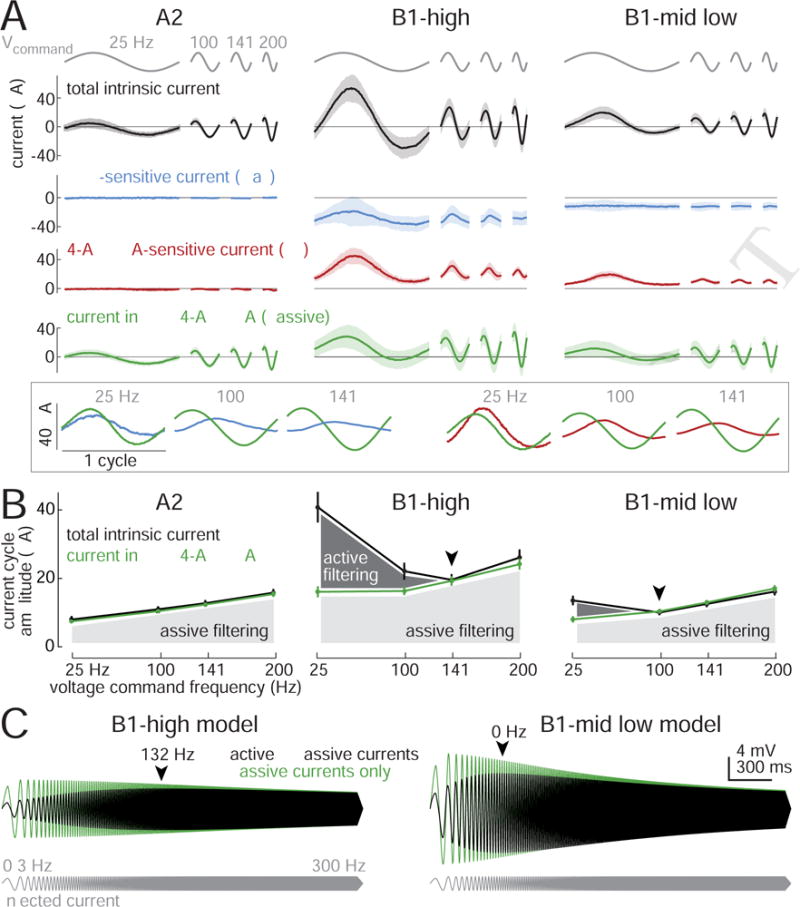
To better understand biophysical mechanisms of mechanosensory processing, we investigated two cell types in the Drosophila brain (A2 and B1 cells) that are postsynaptic to antennal vibration receptors. A2 cells receive excitatory synaptic currents in response to both directions of movement: thus, twice per vibration cycle. The membrane acts as a low-pass filter, so that voltage and spiking mainly track the vibration envelope rather than individual cycles. By contrast, B1 cells are excited by only forward or backward movement, meaning they are sensitive to vibration phase. They receive oscillatory synaptic currents at the stimulus frequency, and they bandpass filter these inputs to favor specific frequencies. Different cells prefer different frequencies, due to differences in their voltage-gated conductances. Both Na+ and K+ conductances suppress low-frequency synaptic inputs, so cells with larger voltage-gated conductances prefer higher frequencies. These results illustrate how membrane properties and voltage-gated conductances can extract distinct stimulus features into parallel channels.
Keywords: AMMC; Johnston's organ; active conductances; depolarization block; electrical synapses; frequency selectivity; mechanosensation; phase selectivity; vibration sensing; voltage-gated ion channels.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Albert JT, Nadrowski B, Göpfert MC. Mechanical signatures of transducer gating in the Drosophila ear. Curr Biol. 2007;17:1000–1006. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
