

SMARCB1 is required for widespread BAF complex-mediated activation of enhancers and bivalent promoters
- PMID: 28945250
- PMCID: PMC5803080
- DOI: 10.1038/ng.3958
SMARCB1 is required for widespread BAF complex-mediated activation of enhancers and bivalent promoters
Abstract
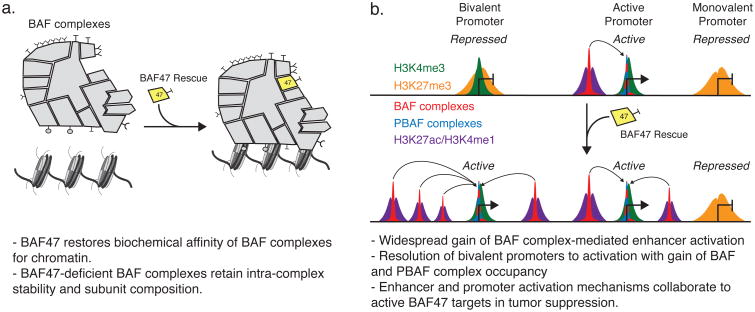
Perturbations to mammalian SWI/SNF (mSWI/SNF or BAF) complexes contribute to more than 20% of human cancers, with driving roles first identified in malignant rhabdoid tumor, an aggressive pediatric cancer characterized by biallelic inactivation of the core BAF complex subunit SMARCB1 (BAF47). However, the mechanism by which this alteration contributes to tumorigenesis remains poorly understood. We find that BAF47 loss destabilizes BAF complexes on chromatin, absent significant changes in complex assembly or integrity. Rescue of BAF47 in BAF47-deficient sarcoma cell lines results in increased genome-wide BAF complex occupancy, facilitating widespread enhancer activation and opposition of Polycomb-mediated repression at bivalent promoters. We demonstrate differential regulation by two distinct mSWI/SNF assemblies, BAF and PBAF complexes, enhancers and promoters, respectively, suggesting that each complex has distinct functions that are perturbed upon BAF47 loss. Our results demonstrate collaborative mechanisms of mSWI/SNF-mediated gene activation, identifying functions that are co-opted or abated to drive human cancers and developmental disorders.
Figures






References
-
- Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell. 2012;150:12–27. - PubMed
-
- Clapier CR, Cairns BR. The biology of chromatin remodeling complexes. Annu Rev Biochem. 2009;78:273–304. - PubMed
-
- Peterson CL, Herskowitz I. Characterization of the yeast SWI1, SWI2, and SWI3 genes, which encode a global activator of transcription. Cell. 1992;68:573–83. - PubMed
-
- Tamkun JW, et al. brahma: a regulator of Drosophila homeotic genes structurally related to the yeast transcriptional activator SNF2/SWI2. Cell. 1992;68:561–72. - PubMed
-
- Kwon H, Imbalzano AN, Khavari PA, Kingston RE, Green MR. Nucleosome disruption and enhancement of activator binding by a human SW1/SNF complex. Nature. 1994;370:477–81. - PubMed
References for Methods
-
- Kadoch C, Copeland RA, Keilhack H. PRC2 and SWI/SNF Chromatin Remodeling Complexes in Health and Disease. Biochemistry. 2016;55:1600–14. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
