

A Modular Receptor Platform To Expand the Sensing Repertoire of Bacteria
- PMID: 28946740
- PMCID: PMC5880506
- DOI: 10.1021/acssynbio.7b00266
A Modular Receptor Platform To Expand the Sensing Repertoire of Bacteria
Abstract
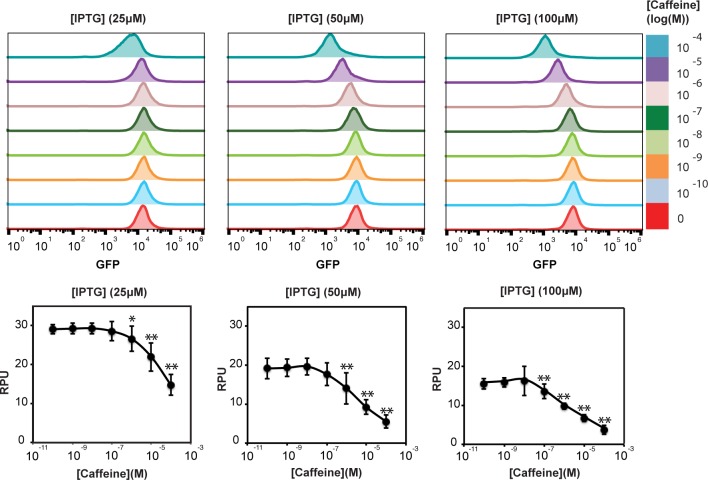
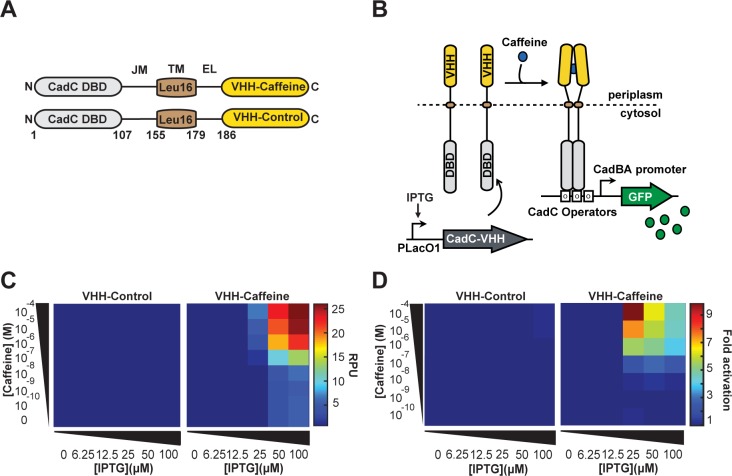
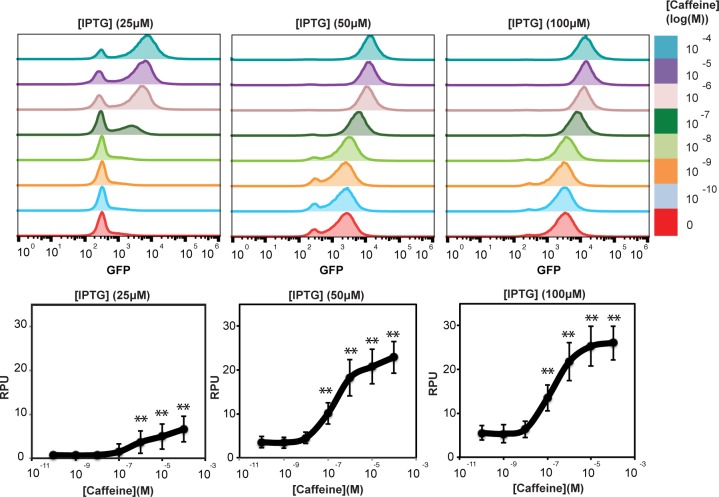
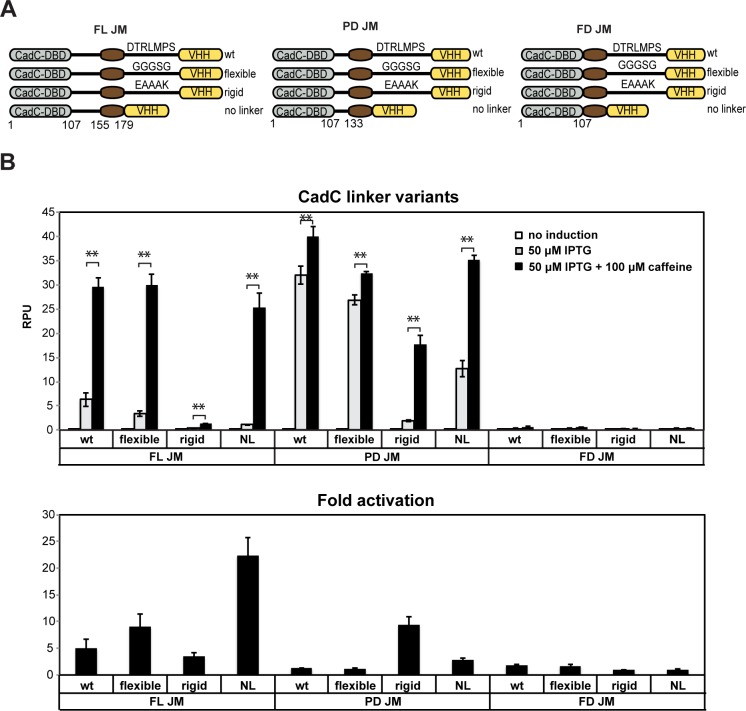
Engineered bacteria promise to revolutionize diagnostics and therapeutics, yet many applications are precluded by the limited number of detectable signals. Here we present a general framework to engineer synthetic receptors enabling bacterial cells to respond to novel ligands. These receptors are activated via ligand-induced dimerization of a single-domain antibody fused to monomeric DNA-binding domains (split-DBDs). Using E. coli as a model system, we engineer both transmembrane and cytosolic receptors using a VHH for ligand detection and demonstrate the scalability of our platform by using the DBDs of two different transcriptional regulators. We provide a method to optimize receptor behavior by finely tuning protein expression levels and optimizing interdomain linker regions. Finally, we show that these receptors can be connected to downstream synthetic gene circuits for further signal processing. The general nature of the split-DBD principle and the versatility of antibody-based detection should support the deployment of these receptors into various hosts to detect ligands for which no receptor is found in nature.
Conflict of interest statement
The authors declare the following competing financial interest(s): We have deposited a patent application on the work presented in the manuscript.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
