

The Photorespiratory Metabolite 2-Phosphoglycolate Regulates Photosynthesis and Starch Accumulation in Arabidopsis
- PMID: 28947491
- PMCID: PMC5774572
- DOI: 10.1105/tpc.17.00256
The Photorespiratory Metabolite 2-Phosphoglycolate Regulates Photosynthesis and Starch Accumulation in Arabidopsis
Abstract
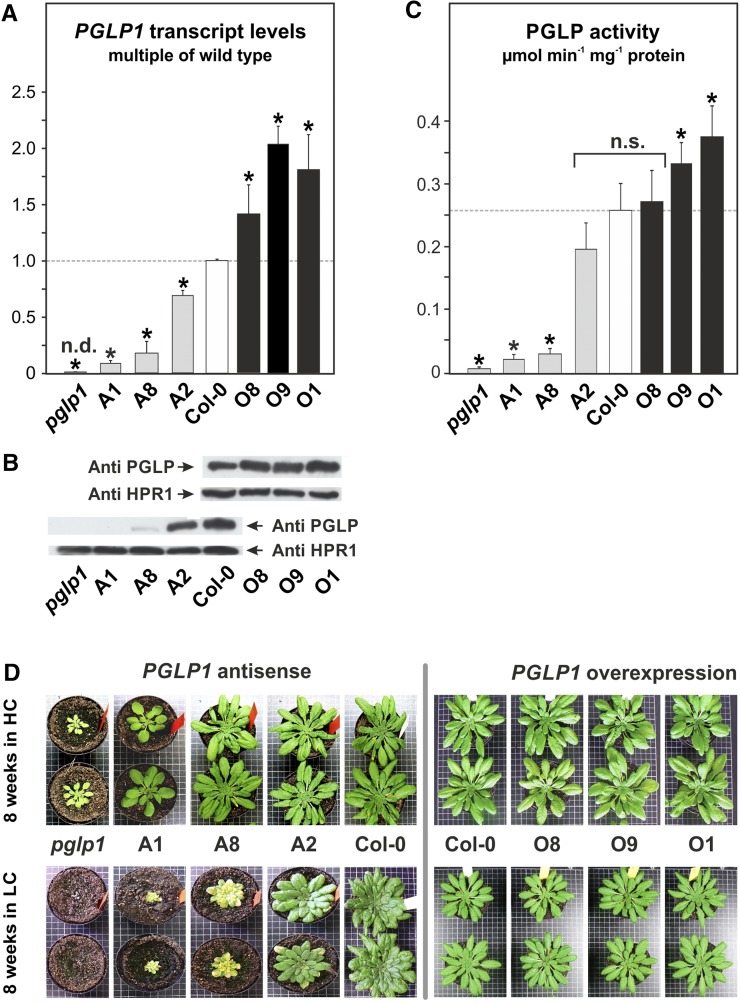
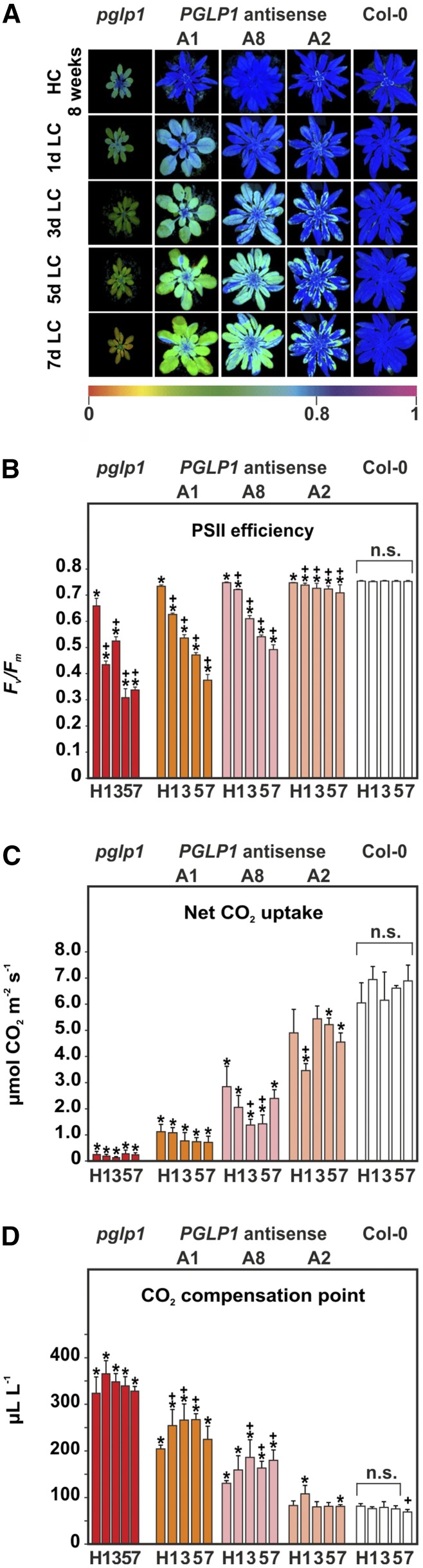
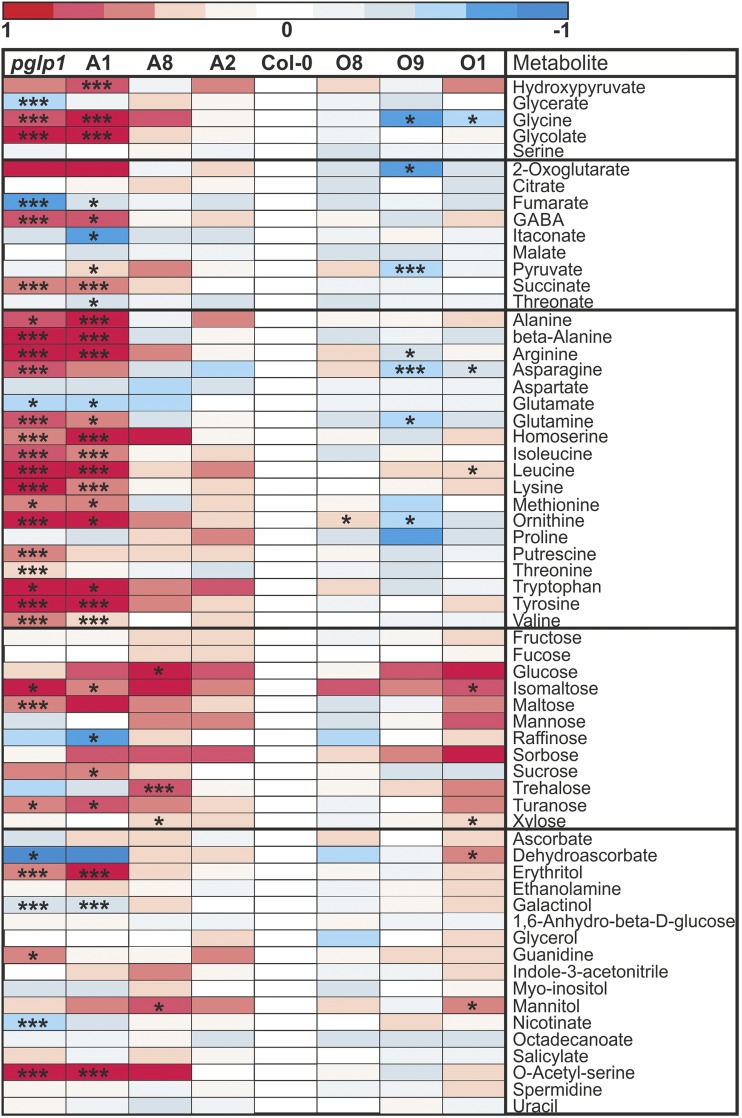
The Calvin-Benson cycle and its photorespiratory repair shunt are in charge of nearly all biological CO2 fixation on Earth. They interact functionally and via shared carbon flow on several levels including common metabolites, transcriptional regulation, and response to environmental changes. 2-Phosphoglycolate (2PG) is one of the shared metabolites and produced in large amounts by oxidative damage of the CO2 acceptor molecule ribulose 1,5-bisphosphate. It was anticipated early on, although never proven, that 2PG could also be a regulatory metabolite that modulates central carbon metabolism by inhibition of triose-phosphate isomerase. Here, we examined this hypothesis using transgenic Arabidopsis thaliana lines with varying activities of the 2PG-degrading enzyme, 2PG phosphatase, and analyzing the impact of this intervention on operation of the Calvin-Benson cycle and other central pathways, leaf carbohydrate metabolism, photosynthetic gas exchange, and growth. Our results demonstrate that 2PG feeds back on the Calvin-Benson cycle. It also alters the allocation of photosynthates between ribulose 1,5-bisphosphate regeneration and starch synthesis. 2PG mechanistically achieves this by inhibiting the Calvin-Benson cycle enzymes triose-phosphate isomerase and sedoheptulose 1,7-bisphosphate phosphatase. We suggest this may represent one of the control loops that sense the ratio of photorespiratory to photosynthetic carbon flux and in turn adjusts stomatal conductance, photosynthetic CO2 and photorespiratory O2 fixation, and starch synthesis in response to changes in the environment.
© 2017 American Society of Plant Biologists. All rights reserved.
Figures


References
-
- Ames B.N. (1966). Assay of inorganic phosphate, total phosphate and phosphatases. In Methods in Enzymology: Complex Carbohydrates, E.F. Neufeld and V. Ginsburg, eds (New York: Academic Press), pp. 115–118.
-
- Anderson L.E. (1971). Chloroplast and cytoplasmic enzymes. II. Pea leaf triose phosphate isomerases. Biochim. Biophys. Acta 235: 237–244. - PubMed
-
- Arrivault S., Guenther M., Ivakov A., Feil R., Vosloh D., van Dongen J.T., Sulpice R., Stitt M. (2009). Use of reverse-phase liquid chromatography, linked to tandem mass spectrometry, to profile the Calvin cycle and other metabolic intermediates in Arabidopsis rosettes at different carbon dioxide concentrations. Plant J. 59: 826–839. - PubMed
-
- Arrivault S., Guenther M., Fry S.C., Fuenfgeld M.M.F.F., Veyel D., Mettler-Altmann T., Stitt M., Lunn J.E. (2015). Synthesis and use of stable-isotope-labeled internal standards for quantification of phosphorylated metabolites by LC-MS/MS. Anal. Chem. 87: 6896–6904. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
