

Rescuing the aberrant sex development of H3K9 demethylase Jmjd1a-deficient mice by modulating H3K9 methylation balance
- PMID: 28949961
- PMCID: PMC5630185
- DOI: 10.1371/journal.pgen.1007034
Rescuing the aberrant sex development of H3K9 demethylase Jmjd1a-deficient mice by modulating H3K9 methylation balance
Abstract
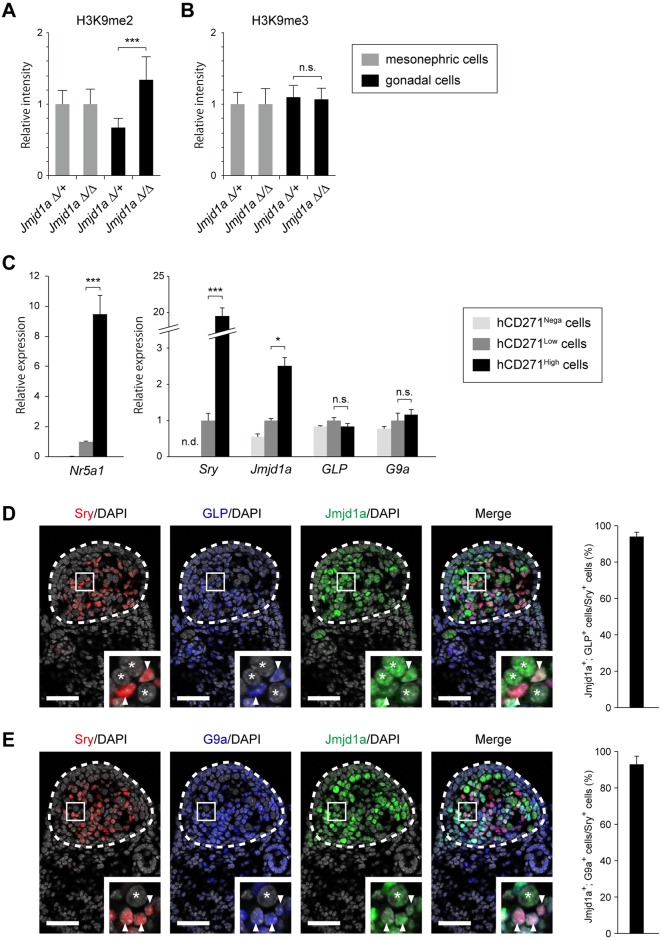
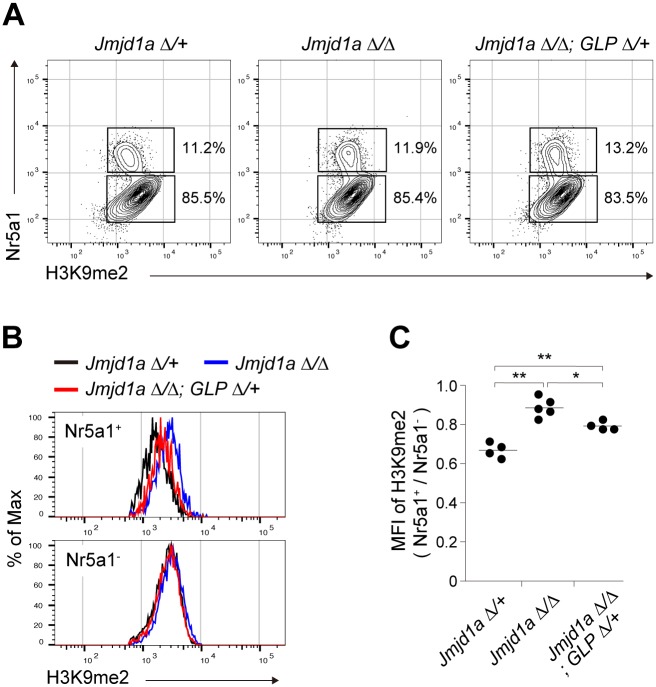
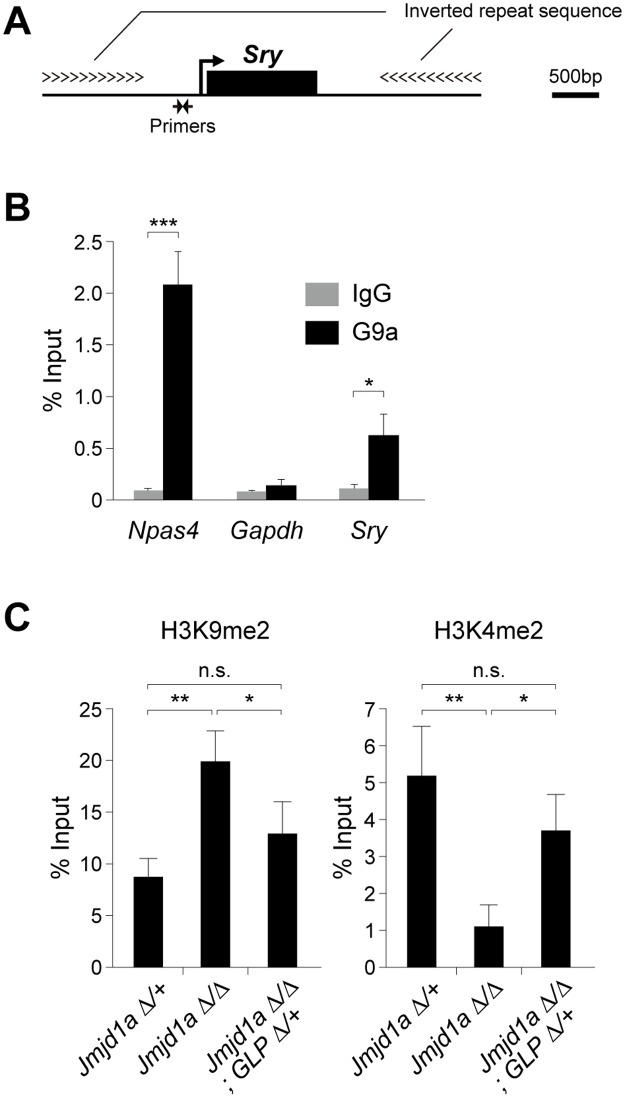
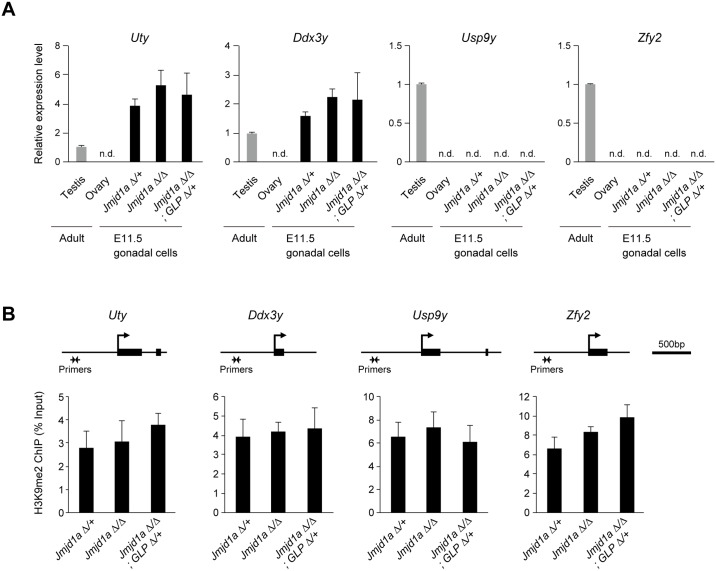
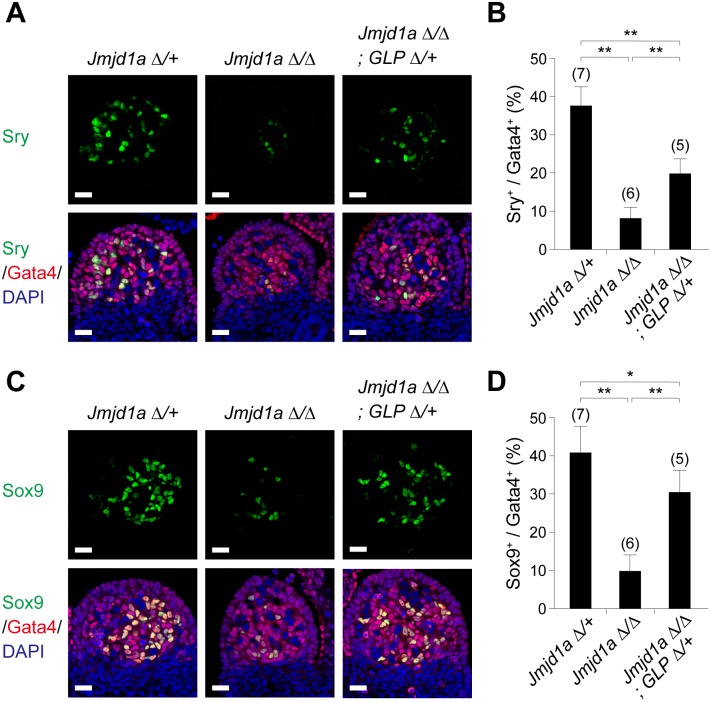
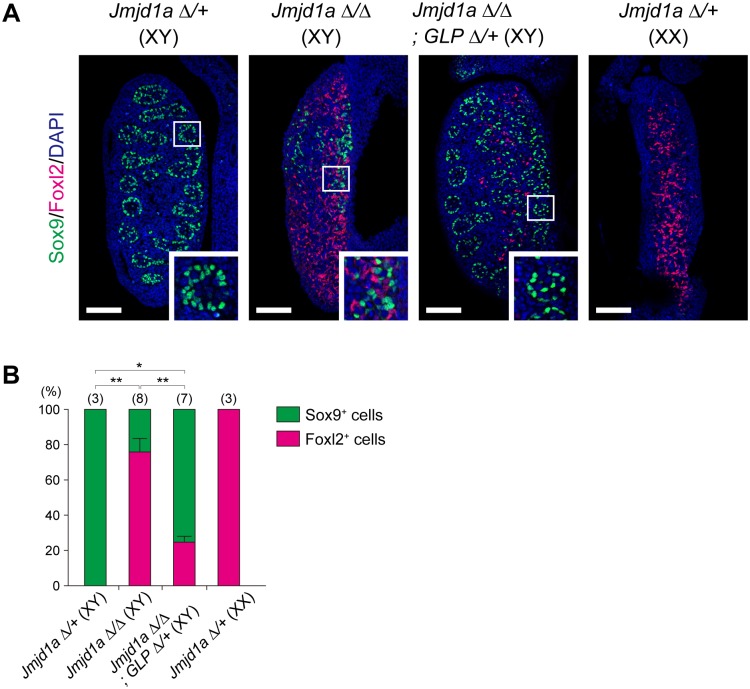
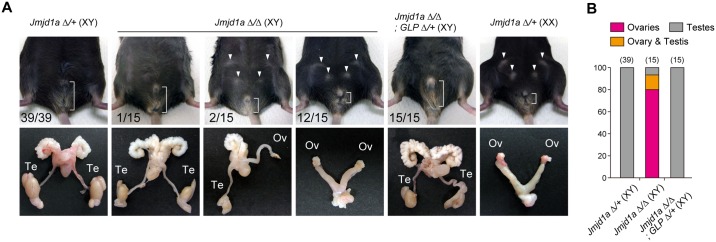
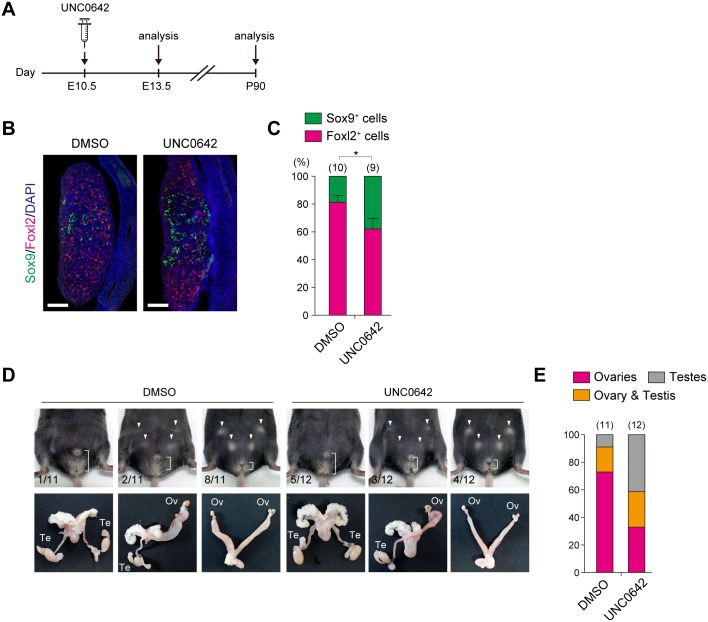
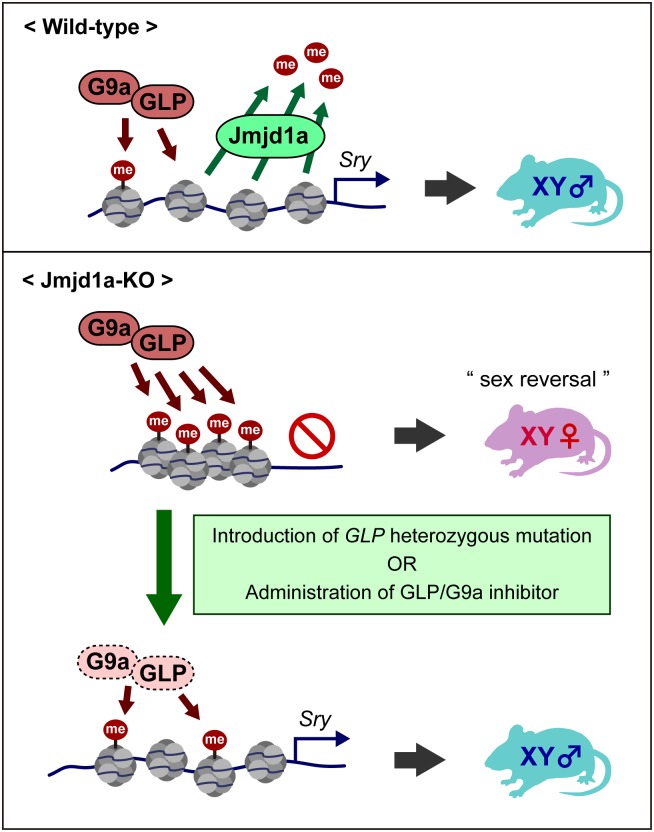
Histone H3 lysine 9 (H3K9) methylation is a hallmark of heterochromatin. H3K9 demethylation is crucial in mouse sex determination; The H3K9 demethylase Jmjd1a deficiency leads to increased H3K9 methylation at the Sry locus in embryonic gonads, thereby compromising Sry expression and causing male-to-female sex reversal. We hypothesized that the H3K9 methylation level at the Sry locus is finely tuned by the balance in activities between the H3K9 demethylase Jmjd1a and an unidentified H3K9 methyltransferase to ensure correct Sry expression. Here we identified the GLP/G9a H3K9 methyltransferase complex as the enzyme catalyzing H3K9 methylation at the Sry locus. Based on this finding, we tried to rescue the sex-reversal phenotype of Jmjd1a-deficient mice by modulating GLP/G9a complex activity. A heterozygous GLP mutation rescued the sex-reversal phenotype of Jmjd1a-deficient mice by restoring Sry expression. The administration of a chemical inhibitor of GLP/G9a enzyme into Jmjd1a-deficient embryos also successfully rescued sex reversal. Our study not only reveals the molecular mechanism underlying the tuning of Sry expression but also provides proof on the principle of therapeutic strategies based on the pharmacological modulation of epigenetic balance.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Greer EL, Shi Y. Histone methylation: a dynamic mark in health, disease and inheritance. Nature reviews Genetics. 2012;13(5):343–57. Epub 2012/04/05. doi: 10.1038/nrg3173 . - DOI - PMC - PubMed
-
- Kuroki S, Matoba S, Akiyoshi M, Matsumura Y, Miyachi H, Mise N, et al. Epigenetic regulation of mouse sex determination by the histone demethylase Jmjd1a. Science. 2013;341(6150):1106–9. Epub 2013/09/07. doi: 10.1126/science.1239864 . - DOI - PubMed
-
- Arrowsmith CH, Bountra C, Fish PV, Lee K, Schapira M. Epigenetic protein families: a new frontier for drug discovery. Nat Rev Drug Discov. 2012;11(5):384–400. doi: 10.1038/nrd3674 . - DOI - PubMed
-
- Rea S, Eisenhaber F, O'Carroll D, Strahl BD, Sun ZW, Schmid M, et al. Regulation of chromatin structure by site-specific histone H3 methyltransferases. Nature. 2000;406(6796):593–9. doi: 10.1038/35020506 . - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
