

Mechanobiology of YAP and TAZ in physiology and disease
- PMID: 28951564
- PMCID: PMC6192510
- DOI: 10.1038/nrm.2017.87
Mechanobiology of YAP and TAZ in physiology and disease
Abstract
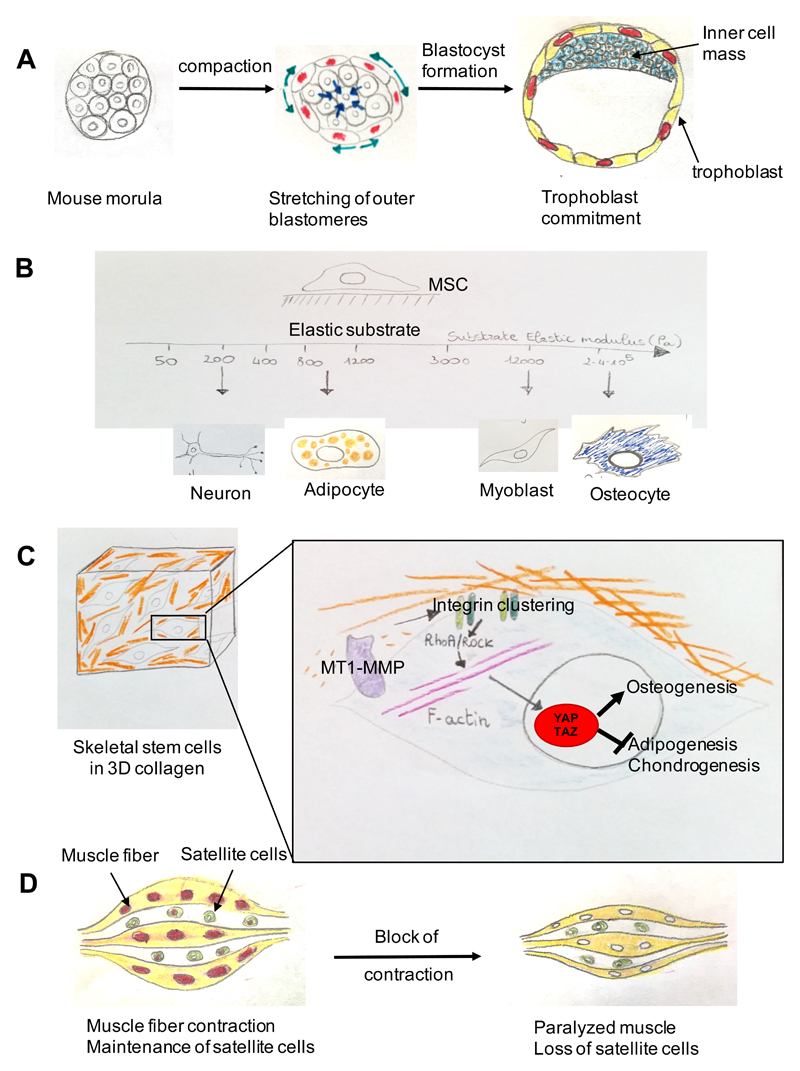
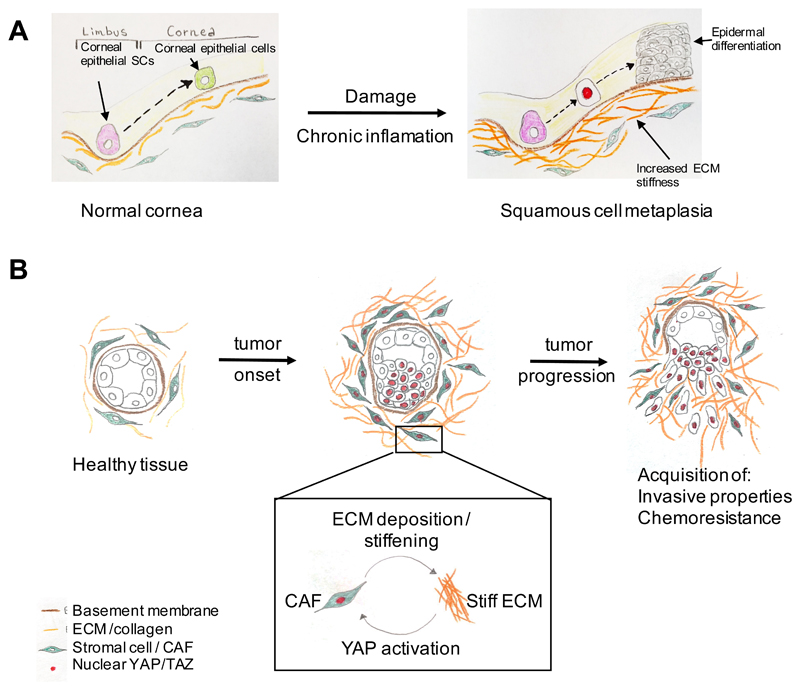
A growing body of evidence suggests that mechanical signals emanating from the cell's microenvironment are fundamental regulators of cell behaviour. Moreover, at the macroscopic scale, the influence of forces, such as the forces generated by blood flow, muscle contraction, gravity and overall tissue rigidity (for example, inside of a tumour lump), is central to our understanding of physiology and disease pathogenesis. Still, how mechanical cues are sensed and transduced at the molecular level to regulate gene expression has long remained enigmatic. The identification of the transcription factors YAP and TAZ as mechanotransducers started to fill this gap. YAP and TAZ read a broad range of mechanical cues, from shear stress to cell shape and extracellular matrix rigidity, and translate them into cell-specific transcriptional programmes. YAP and TAZ mechanotransduction is critical for driving stem cell behaviour and regeneration, and it sheds new light on the mechanisms by which aberrant cell mechanics is instrumental for the onset of multiple diseases, such as atherosclerosis, fibrosis, pulmonary hypertension, inflammation, muscular dystrophy and cancer.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
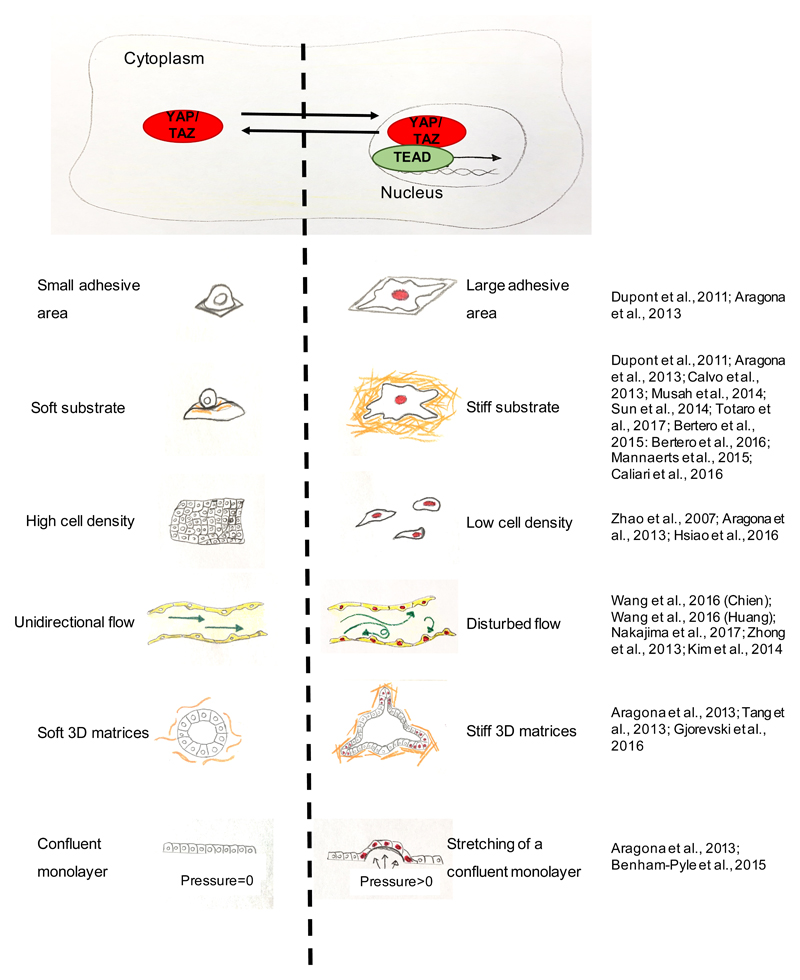
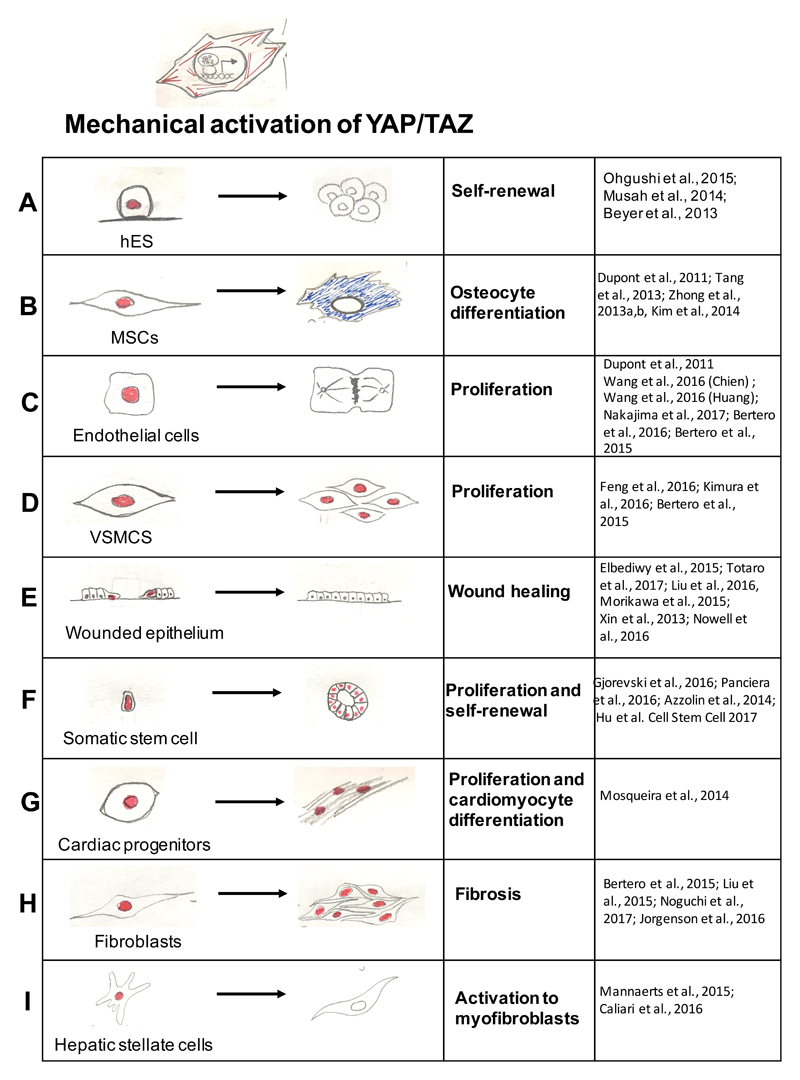
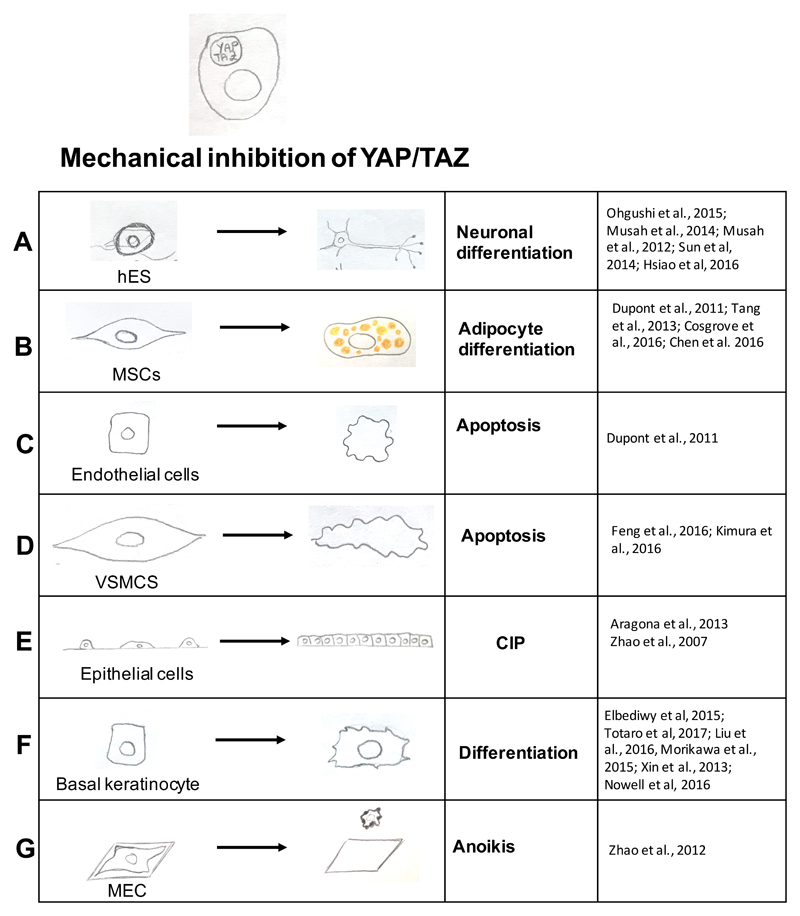
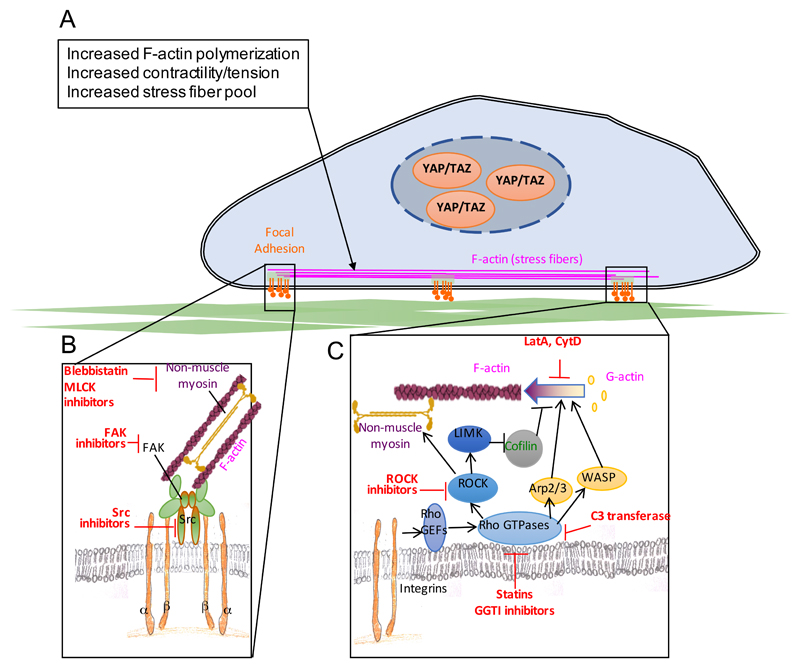
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
