

Dynamic patterning by the Drosophila pair-rule network reconciles long-germ and short-germ segmentation
- PMID: 28953896
- PMCID: PMC5633203
- DOI: 10.1371/journal.pbio.2002439
Dynamic patterning by the Drosophila pair-rule network reconciles long-germ and short-germ segmentation
Abstract
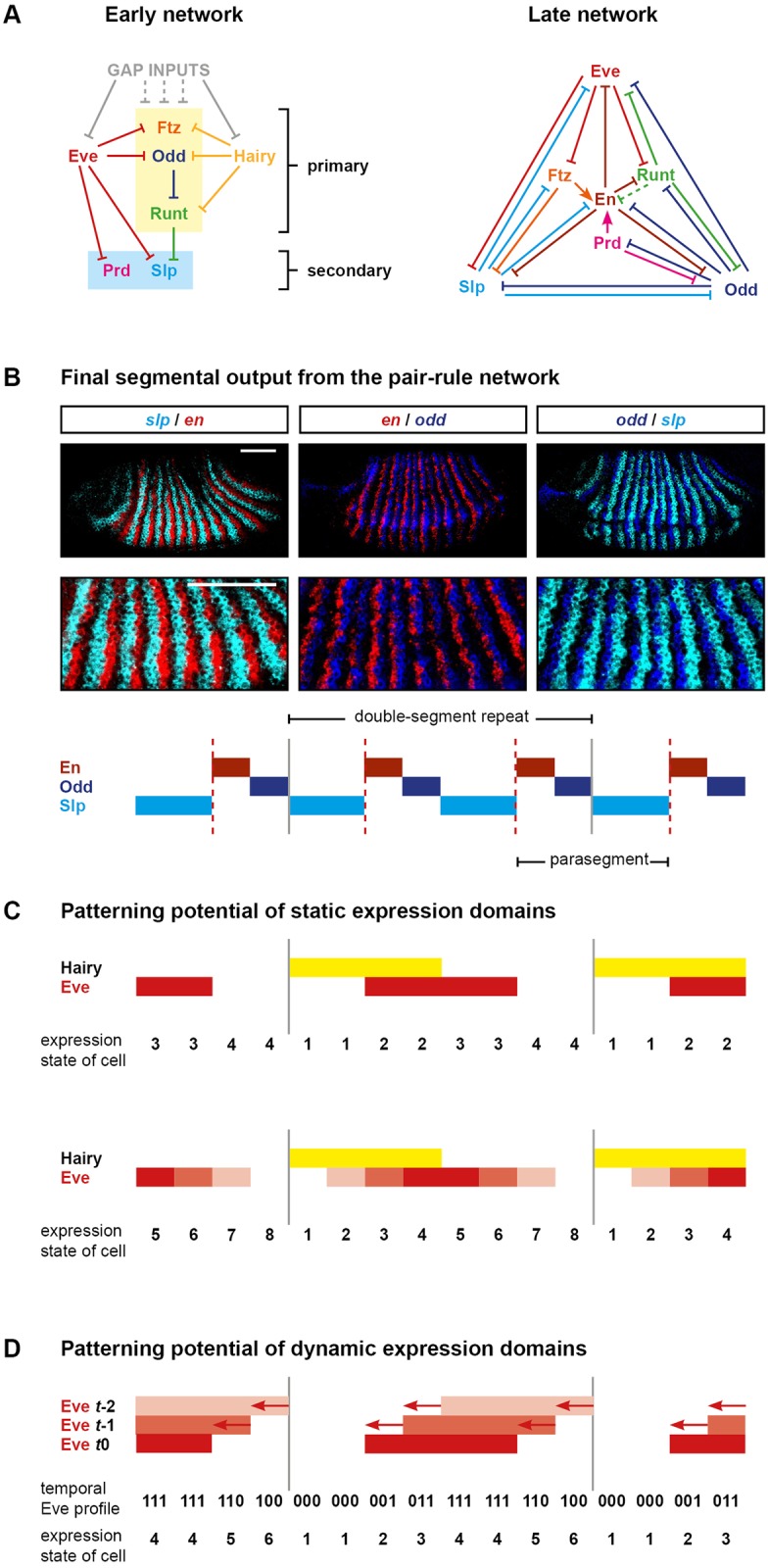
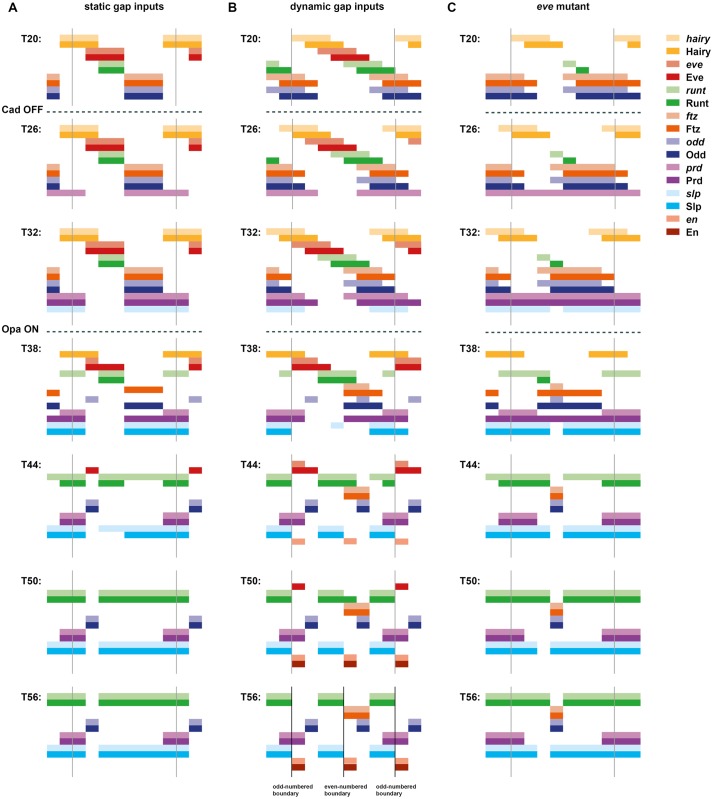
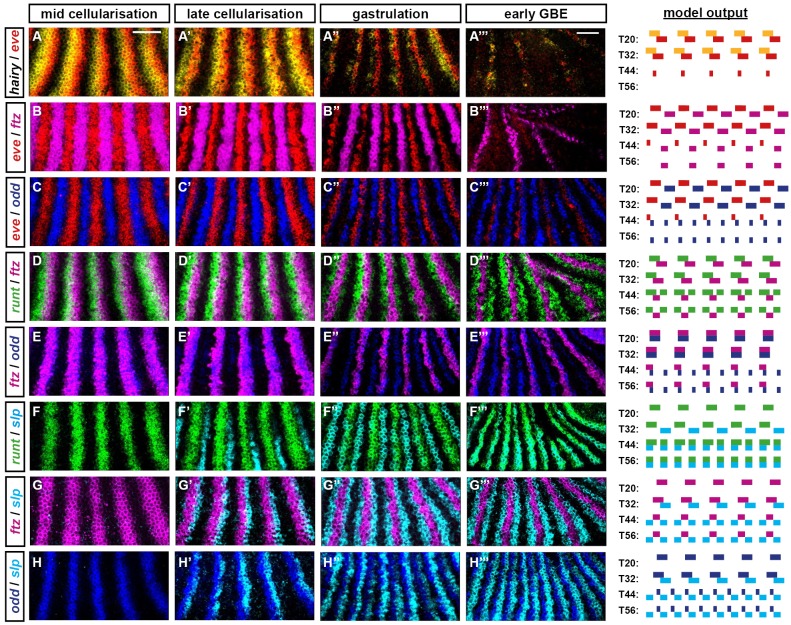
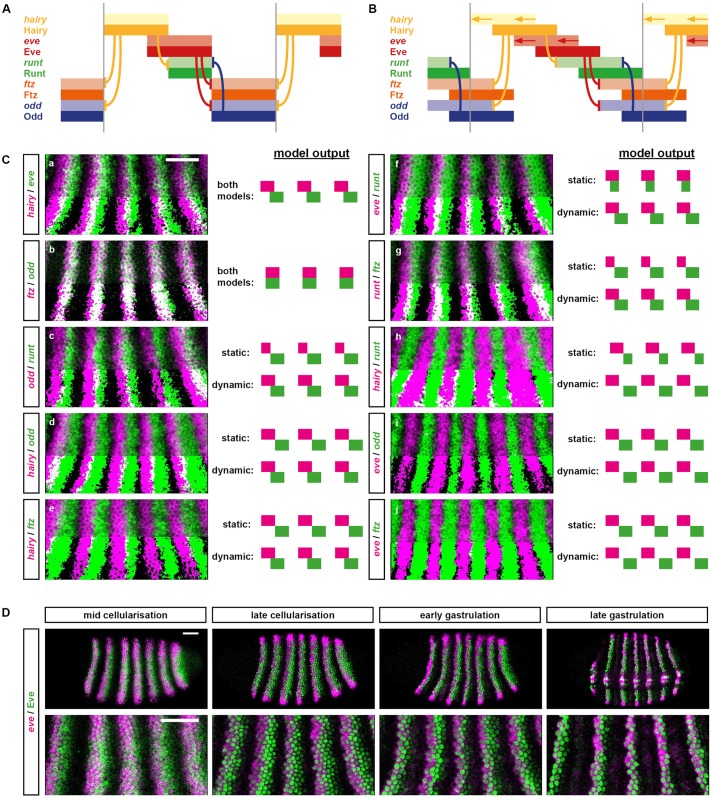
Drosophila segmentation is a well-established paradigm for developmental pattern formation. However, the later stages of segment patterning, regulated by the "pair-rule" genes, are still not well understood at the system level. Building on established genetic interactions, I construct a logical model of the Drosophila pair-rule system that takes into account the demonstrated stage-specific architecture of the pair-rule gene network. Simulation of this model can accurately recapitulate the observed spatiotemporal expression of the pair-rule genes, but only when the system is provided with dynamic "gap" inputs. This result suggests that dynamic shifts of pair-rule stripes are essential for segment patterning in the trunk and provides a functional role for observed posterior-to-anterior gap domain shifts that occur during cellularisation. The model also suggests revised patterning mechanisms for the parasegment boundaries and explains the aetiology of the even-skipped null mutant phenotype. Strikingly, a slightly modified version of the model is able to pattern segments in either simultaneous or sequential modes, depending only on initial conditions. This suggests that fundamentally similar mechanisms may underlie segmentation in short-germ and long-germ arthropods.
Conflict of interest statement
The author has declared that no competing interests exist.
Figures
Similar articles
-
Expression of pair rule gene orthologs in the blastoderm of a myriapod: evidence for pair rule-like mechanisms?BMC Dev Biol. 2012 May 17;12:15. doi: 10.1186/1471-213X-12-15. BMC Dev Biol. 2012. PMID: 22595029 Free PMC article.
-
A systematic analysis of the gap gene system in the moth midge Clogmia albipunctata.Dev Biol. 2010 Aug 1;344(1):306-18. doi: 10.1016/j.ydbio.2010.04.019. Epub 2010 Apr 28. Dev Biol. 2010. PMID: 20433825
-
Shifting roles of Drosophila pair-rule gene orthologs: segmental expression and function in the milkweed bug Oncopeltus fasciatus.Development. 2019 Sep 10;146(17):dev181453. doi: 10.1242/dev.181453. Development. 2019. PMID: 31444220 Free PMC article.
-
Forces directing germ-band extension in Drosophila embryos.Mech Dev. 2017 Apr;144(Pt A):11-22. doi: 10.1016/j.mod.2016.12.001. Epub 2016 Dec 22. Mech Dev. 2017. PMID: 28013027 Review.
-
The gap gene network.Cell Mol Life Sci. 2011 Jan;68(2):243-74. doi: 10.1007/s00018-010-0536-y. Epub 2010 Oct 8. Cell Mol Life Sci. 2011. PMID: 20927566 Free PMC article. Review.
Cited by
-
The Evolution of Sox Gene Repertoires and Regulation of Segmentation in Arachnids.Mol Biol Evol. 2021 Jul 29;38(8):3153-3169. doi: 10.1093/molbev/msab088. Mol Biol Evol. 2021. PMID: 33755150 Free PMC article.
-
Developmental Bias and Evolution: A Regulatory Network Perspective.Genetics. 2018 Aug;209(4):949-966. doi: 10.1534/genetics.118.300995. Genetics. 2018. PMID: 30049818 Free PMC article. Review.
-
Striped Expression of Leucine-Rich Repeat Proteins Coordinates Cell Intercalation and Compartment Boundary Formation in the Early Drosophila Embryo.Symmetry (Basel). 2023 Aug;15(8):1490. doi: 10.3390/sym15081490. Epub 2023 Jul 27. Symmetry (Basel). 2023. PMID: 38650964 Free PMC article.
-
Quantitative analysis reveals genotype- and domain- specific differences between mRNA and protein expression of segmentation genes in Drosophila.Dev Biol. 2019 Apr 1;448(1):48-58. doi: 10.1016/j.ydbio.2019.01.006. Epub 2019 Jan 7. Dev Biol. 2019. PMID: 30629954 Free PMC article.
-
A re-inducible gap gene cascade patterns the anterior-posterior axis of insects in a threshold-free fashion.Elife. 2018 Dec 20;7:e41208. doi: 10.7554/eLife.41208. Elife. 2018. PMID: 30570485 Free PMC article.
References
-
- Akam M. The molecular basis for metameric pattern in the Drosophila embryo. Development. 1987;101:1–22. - PubMed
-
- Nasiadka A, Dietrich BH, Krause HM. Anterior—posterior patterning in the Drosophila embryo. Adv Dev Biol Biochem. 2002;12:155–204.
-
- Nüsslein-Volhard C, Wieschaus E. Mutations affecting segment number and polarity in Drosophila. Nature. 1980;287:795–801. - PubMed
-
- Wieschaus E, Nüsslein-Volhard C. The Heidelberg Screen for Pattern Mutants of Drosophila : A Personal Account. Annu Rev Cell Dev Biol. 2016;32:1–46. doi: 10.1146/annurev-cellbio-113015-023138 - DOI - PubMed
-
- Arnosti DN. Analysis and function of transcriptional regulatory elements: insights from Drosophila. Annu Rev Entomol. 2003;48:579–602. doi: 10.1146/annurev.ento.48.091801.112749 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
