

Hippocampal GABAergic Inhibitory Interneurons
- PMID: 28954853
- PMCID: PMC6151493
- DOI: 10.1152/physrev.00007.2017
Hippocampal GABAergic Inhibitory Interneurons
Abstract
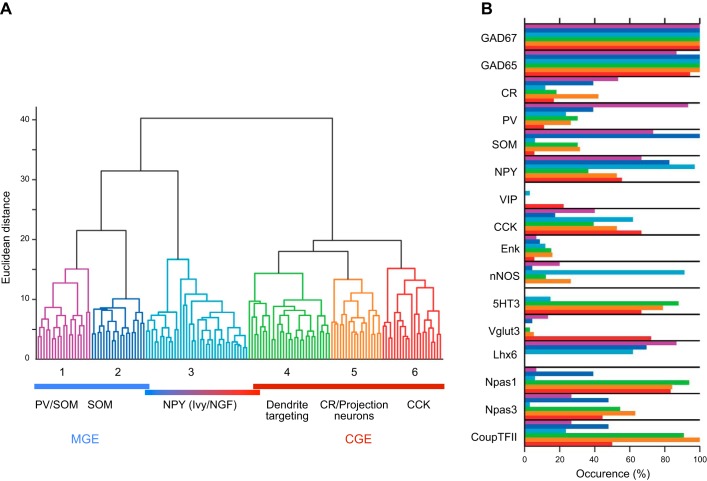
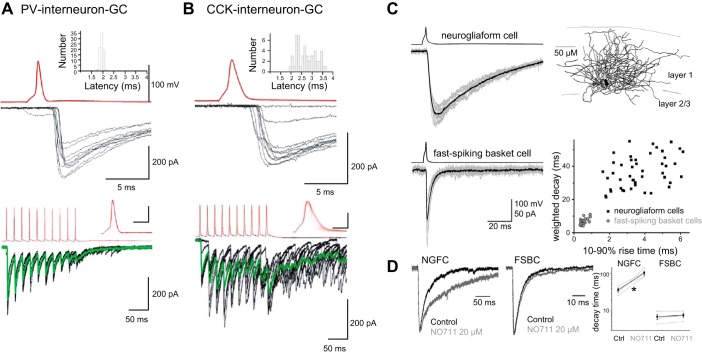
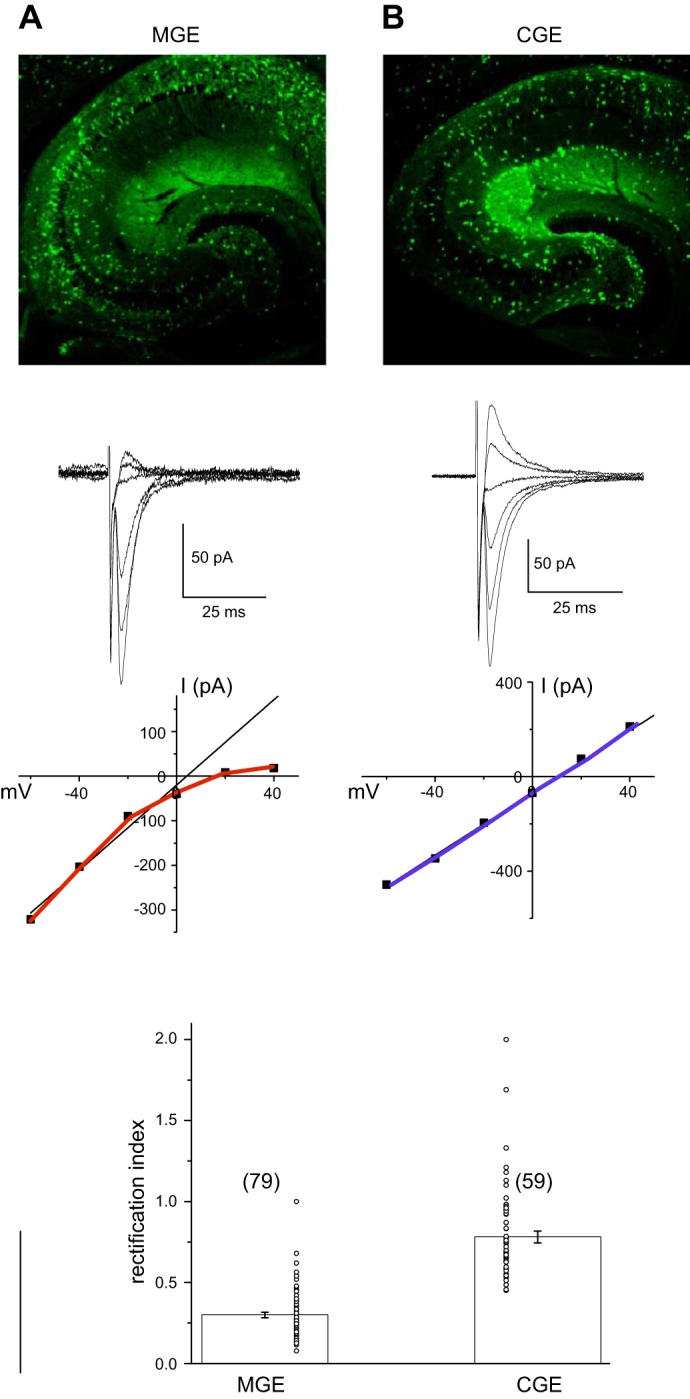
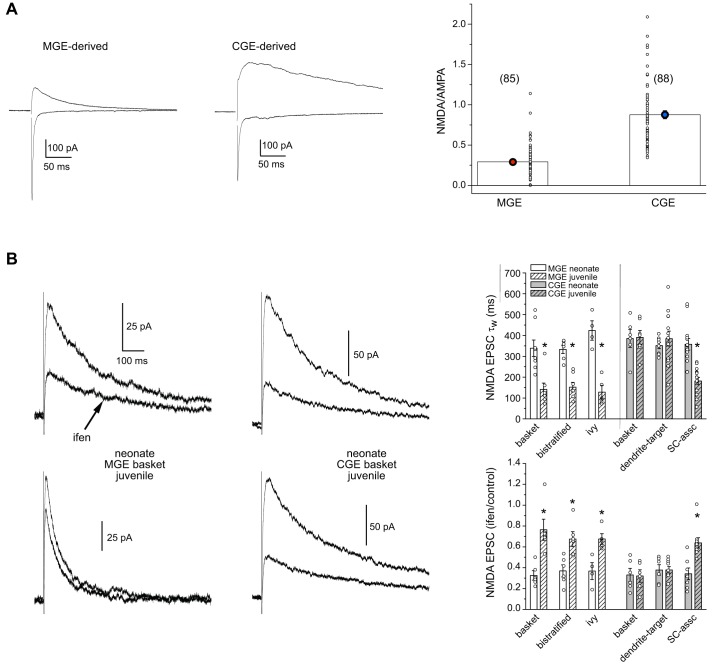
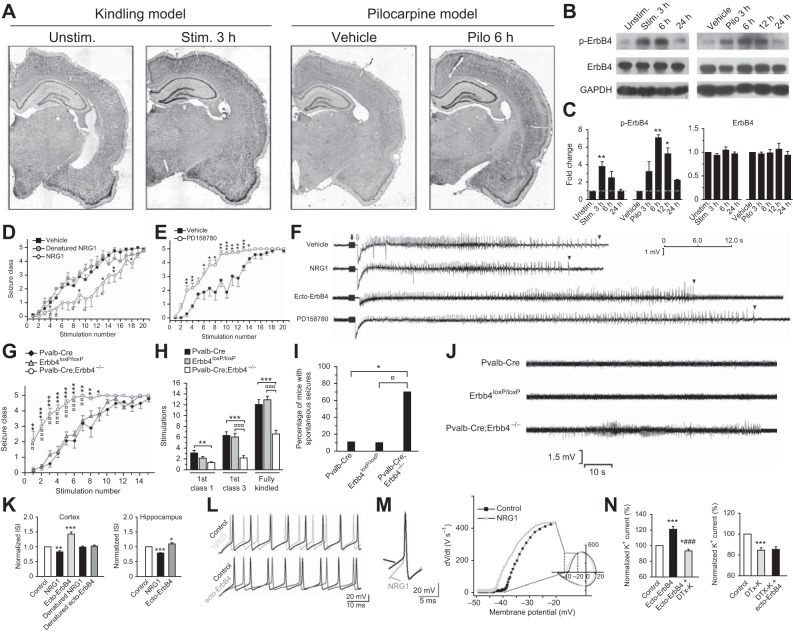
In the hippocampus GABAergic local circuit inhibitory interneurons represent only ~10-15% of the total neuronal population; however, their remarkable anatomical and physiological diversity allows them to regulate virtually all aspects of cellular and circuit function. Here we provide an overview of the current state of the field of interneuron research, focusing largely on the hippocampus. We discuss recent advances related to the various cell types, including their development and maturation, expression of subtype-specific voltage- and ligand-gated channels, and their roles in network oscillations. We also discuss recent technological advances and approaches that have permitted high-resolution, subtype-specific examination of their roles in numerous neural circuit disorders and the emerging therapeutic strategies to ameliorate such pathophysiological conditions. The ultimate goal of this review is not only to provide a touchstone for the current state of the field, but to help pave the way for future research by highlighting where gaps in our knowledge exist and how a complete appreciation of their roles will aid in future therapeutic strategies.
Copyright © 2017 the American Physiological Society.
Figures
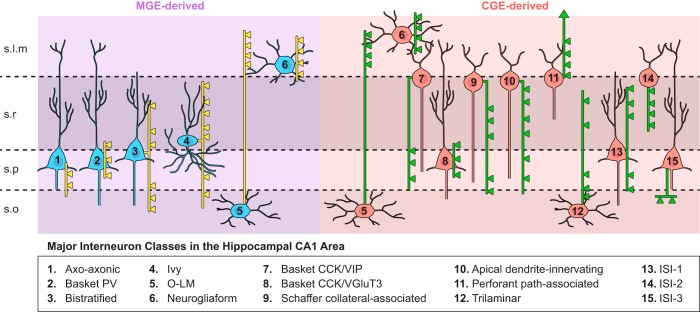
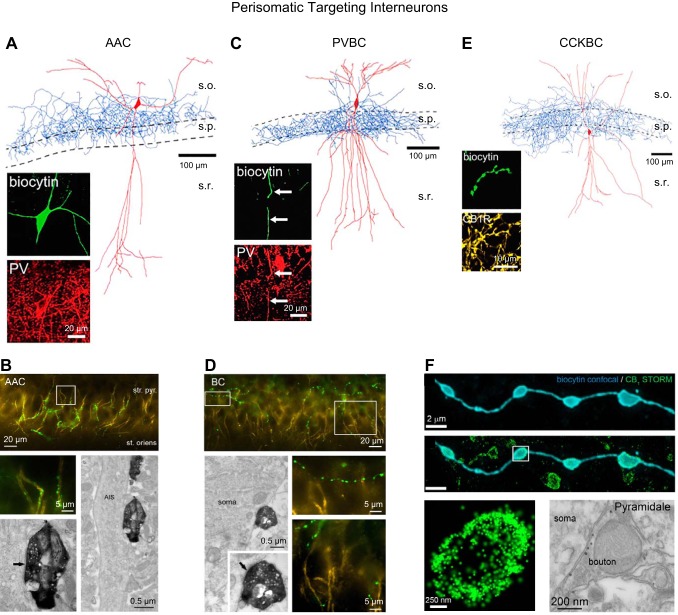
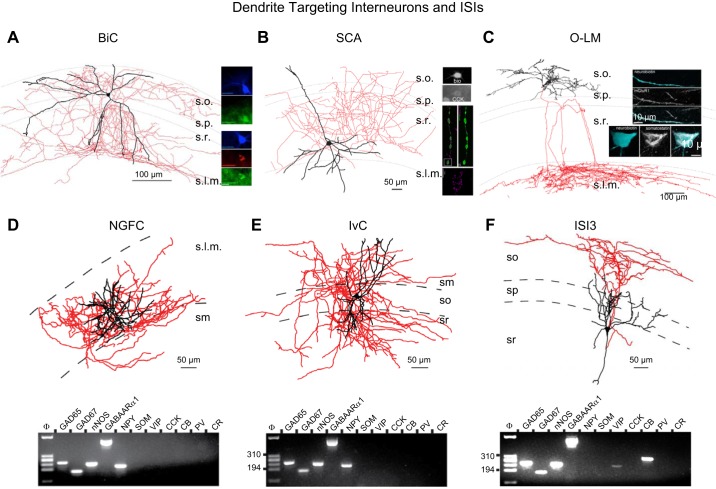

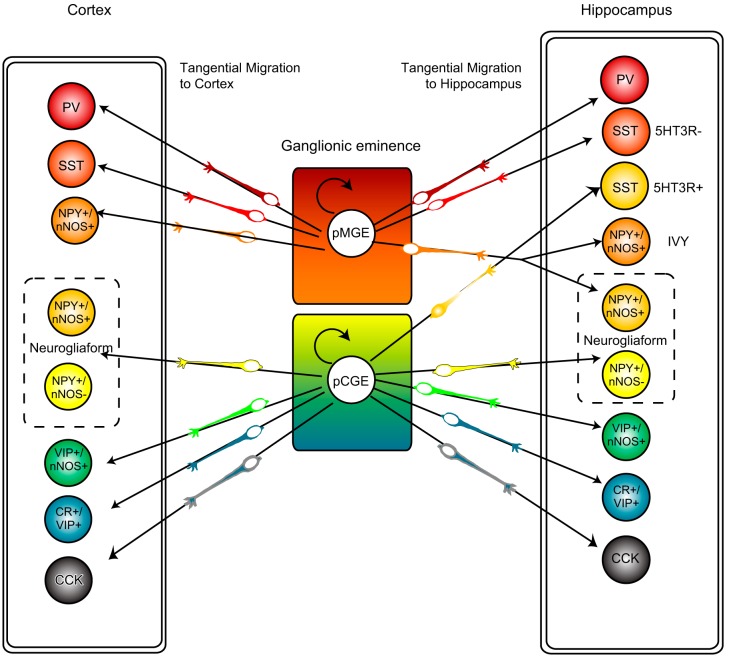
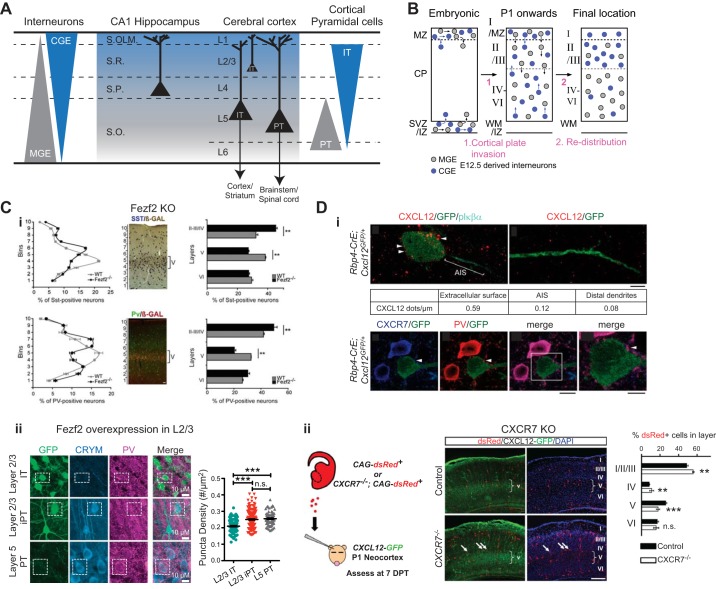
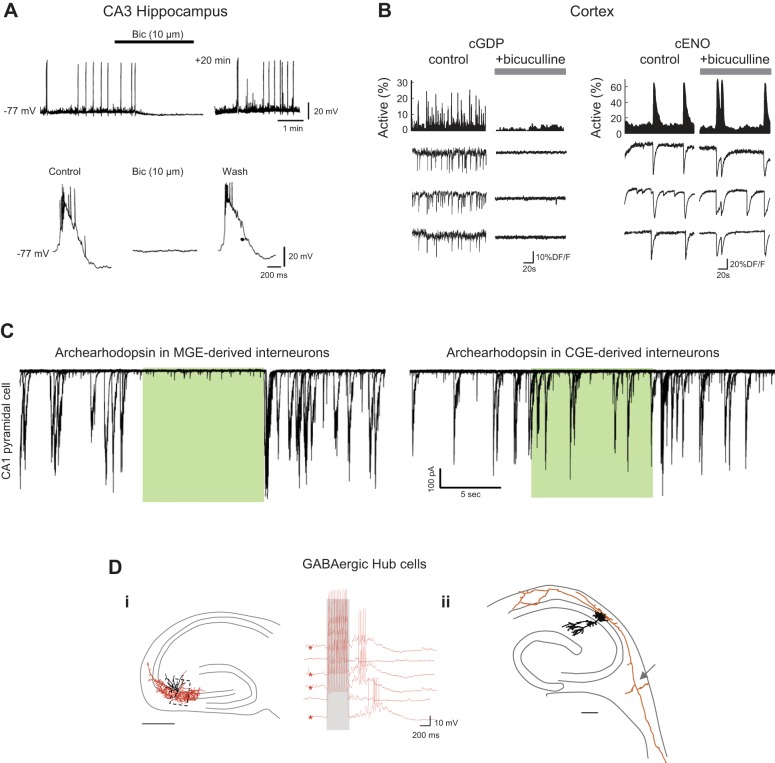


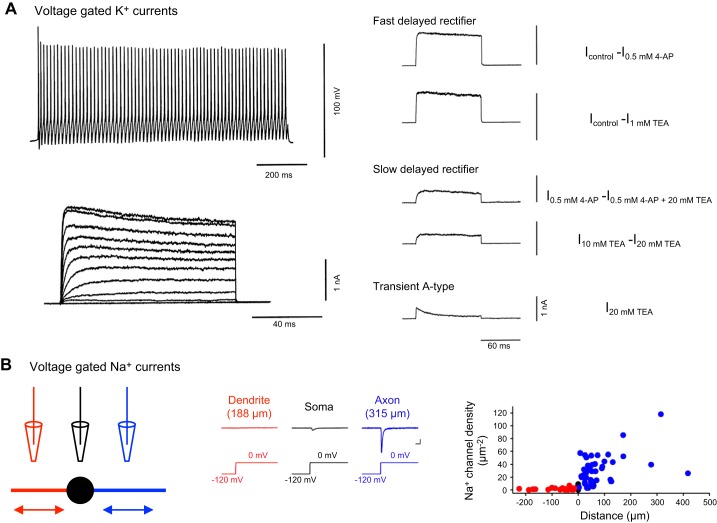
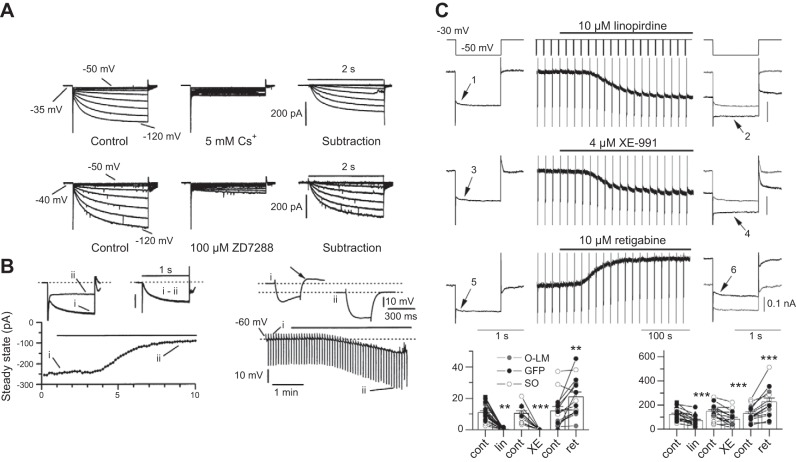
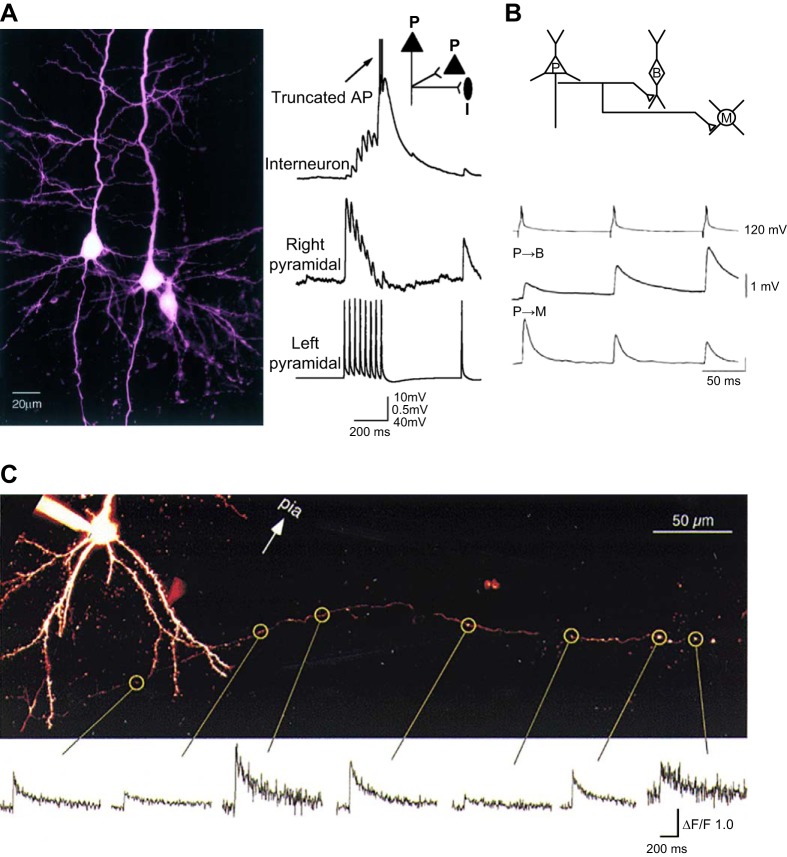
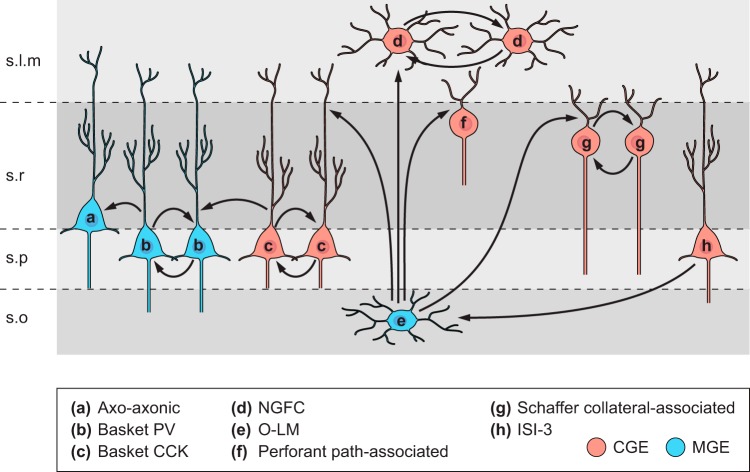
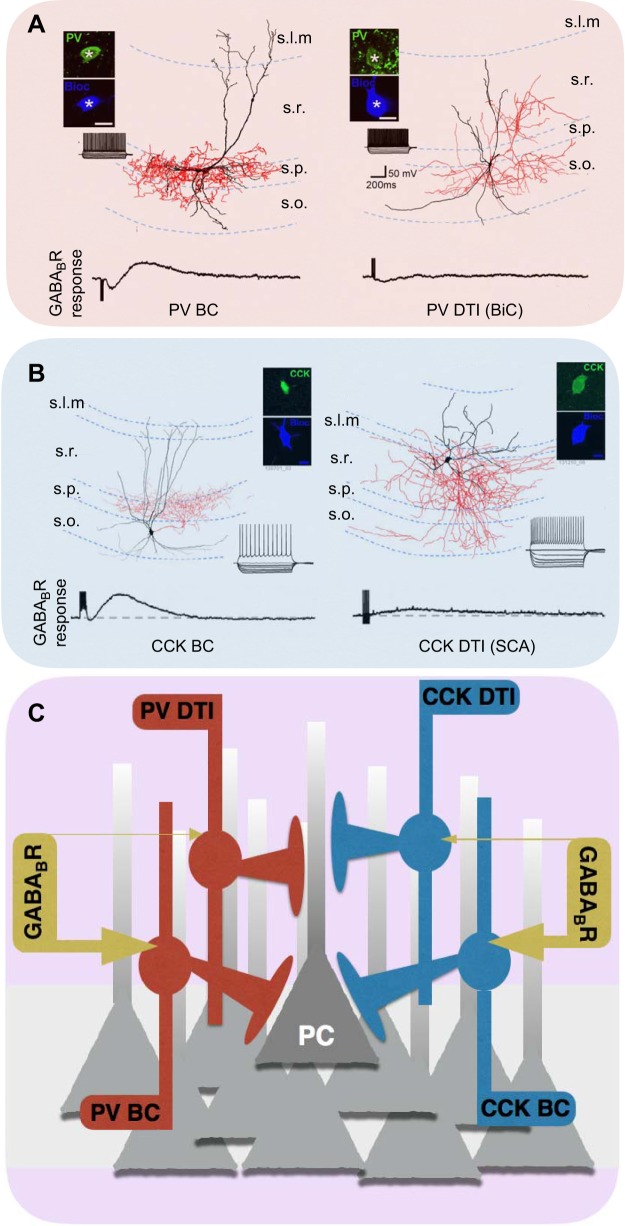
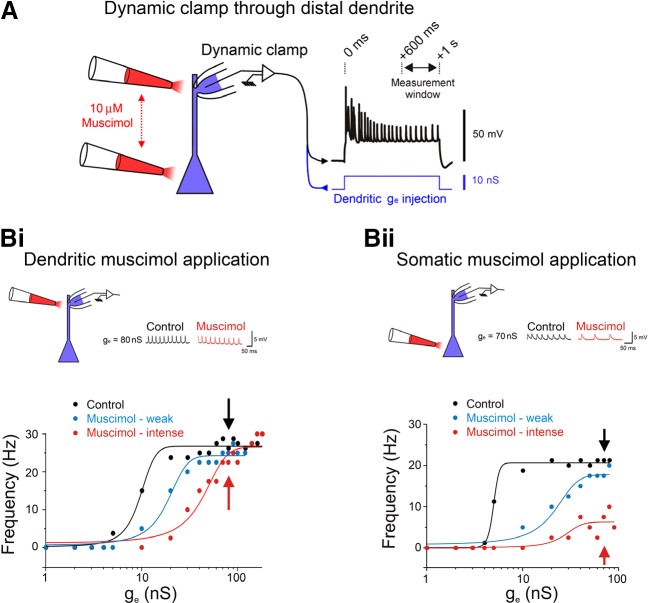
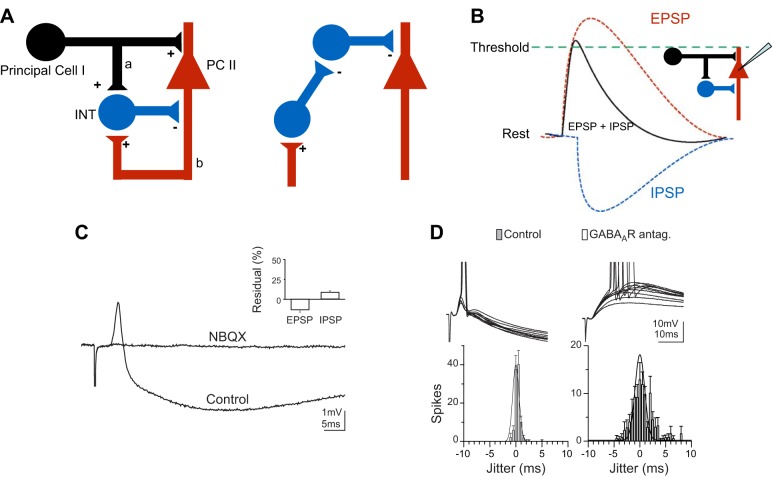
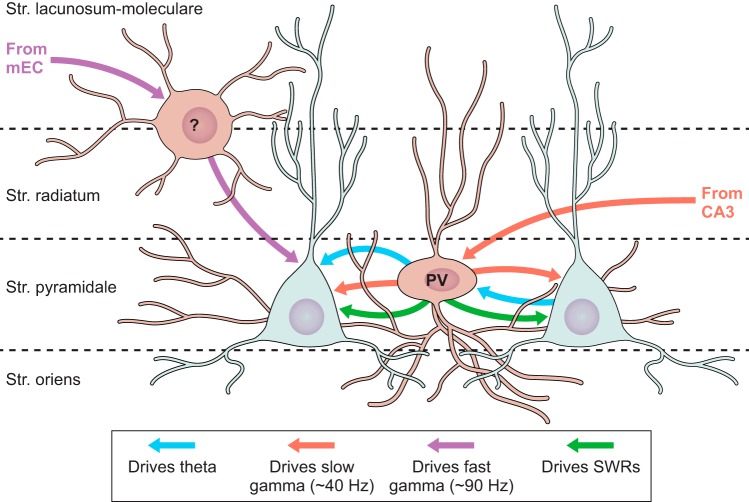

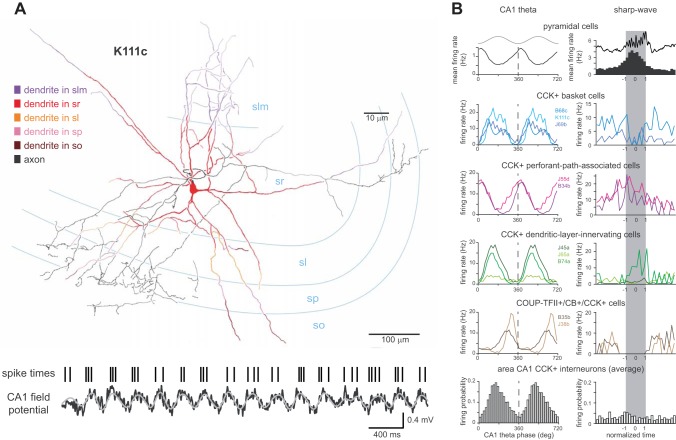
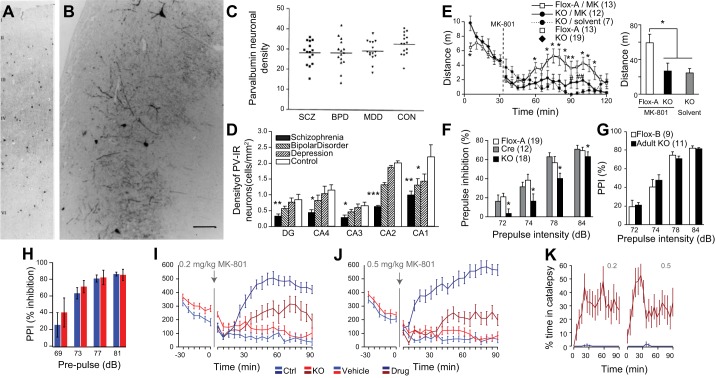
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources