

RNAi-based treatment of chronically infected patients and chimpanzees reveals that integrated hepatitis B virus DNA is a source of HBsAg
- PMID: 28954926
- PMCID: PMC5830187
- DOI: 10.1126/scitranslmed.aan0241
RNAi-based treatment of chronically infected patients and chimpanzees reveals that integrated hepatitis B virus DNA is a source of HBsAg
Abstract
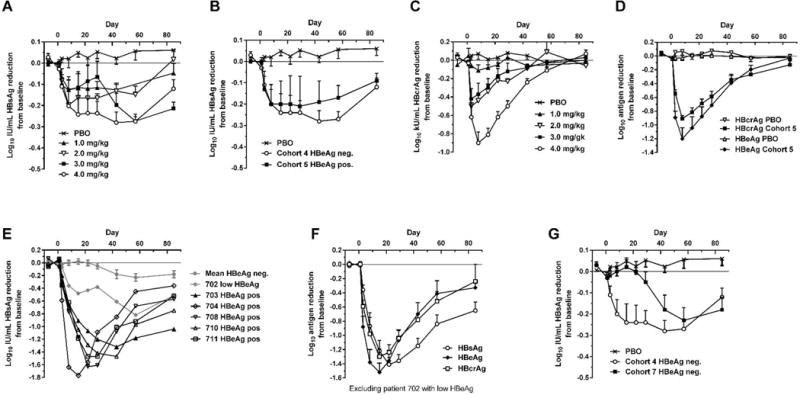
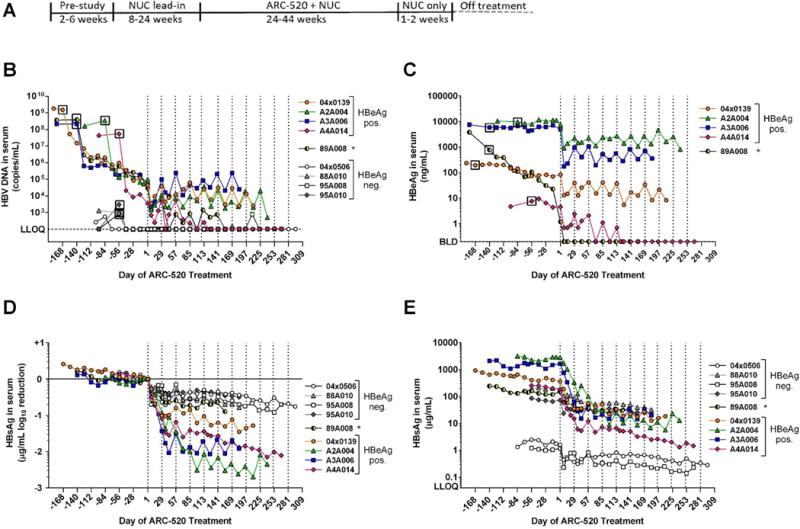
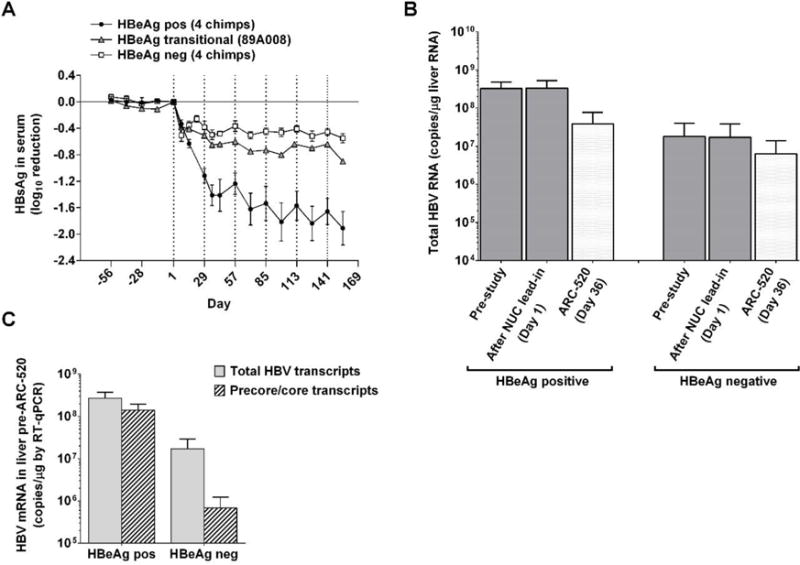
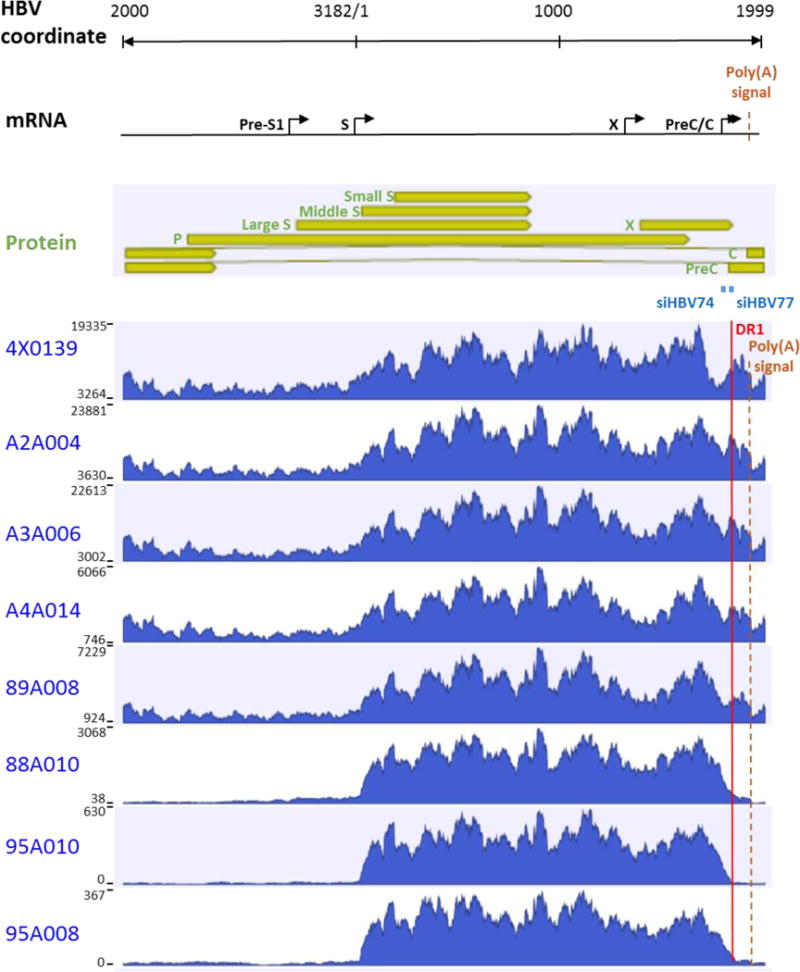
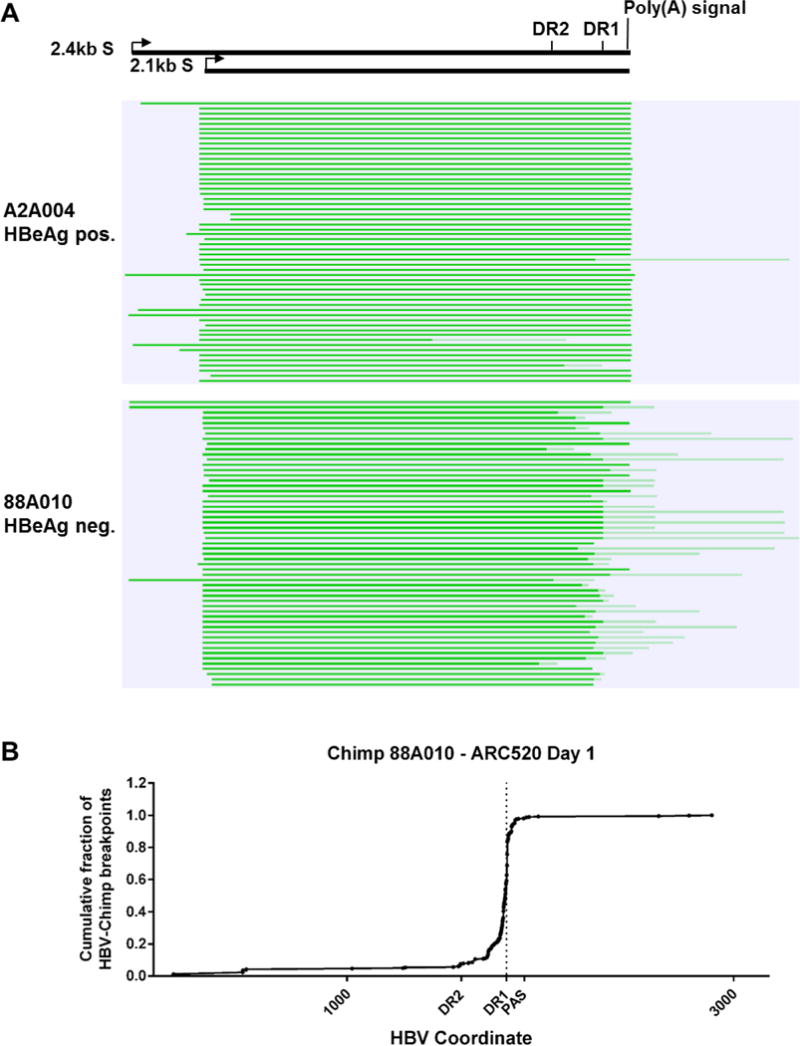
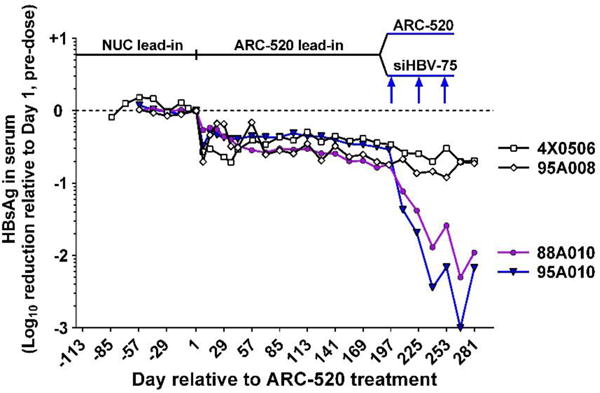
Chronic hepatitis B virus (HBV) infection is a major health concern worldwide, frequently leading to liver cirrhosis, liver failure, and hepatocellular carcinoma. Evidence suggests that high viral antigen load may play a role in chronicity. Production of viral proteins is thought to depend on transcription of viral covalently closed circular DNA (cccDNA). In a human clinical trial with an RNA interference (RNAi)-based therapeutic targeting HBV transcripts, ARC-520, HBV S antigen (HBsAg) was strongly reduced in treatment-naïve patients positive for HBV e antigen (HBeAg) but was reduced significantly less in patients who were HBeAg-negative or had received long-term therapy with nucleos(t)ide viral replication inhibitors (NUCs). HBeAg positivity is associated with greater disease risk that may be moderately reduced upon HBeAg loss. The molecular basis for this unexpected differential response was investigated in chimpanzees chronically infected with HBV. Several lines of evidence demonstrated that HBsAg was expressed not only from the episomal cccDNA minichromosome but also from transcripts arising from HBV DNA integrated into the host genome, which was the dominant source in HBeAg-negative chimpanzees. Many of the integrants detected in chimpanzees lacked target sites for the small interfering RNAs in ARC-520, explaining the reduced response in HBeAg-negative chimpanzees and, by extension, in HBeAg-negative patients. Our results uncover a heretofore underrecognized source of HBsAg that may represent a strategy adopted by HBV to maintain chronicity in the presence of host immunosurveillance. These results could alter trial design and endpoint expectations of new therapies for chronic HBV.
Copyright © 2017 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
C.I.W., D.L.L, Z.X., B.D.G., T.S., J.H., and S.B.K. are or were employees of Arrowhead Pharmaceuticals. R.G.G., S.A.L., C.L.L., C.F. and J.Y.N.L. receive compensation from Arrowhead. M.F.Y., H.Y.C., R.E.L. and D. C. performed work under contract with Arrowhead.
Figures
Comment in
-
Hepatitis: No cure for hepatitis B and D without targeting integrated viral DNA?Nat Rev Gastroenterol Hepatol. 2018 Apr;15(4):195-196. doi: 10.1038/nrgastro.2017.185. Epub 2018 Jan 31. Nat Rev Gastroenterol Hepatol. 2018. PMID: 29382951 No abstract available.
References
-
- Yuen MF, Ahn SH, Chen DS, Chen PJ, Dusheiko GM, Hou JL, Maddrey WC, Mizokami M, Seto WK, Zoulim F, Lai CL. Chronic Hepatitis B Virus Infection: Disease Revisit and Management Recommendations. J Clin Gastroenterol. 2016;50:286–294. - PubMed
-
- Beasley RP, Hwang LY, Lin CC, Chien CS. Hepatocellular carcinoma and hepatitis B virus. A prospective study of 22 707 men in Taiwan. Lancet. 1981;2:1129–1133. - PubMed
-
- Bock CT, Schwinn S, Locarnini S, Fyfe J, Manns MP, Trautwein C, Zentgraf H. Structural organization of the hepatitis B virus minichromosome. J Mol Biol. 2001;307:183–196. - PubMed
-
- Jiang Z, Jhunjhunwala S, Liu J, Haverty PM, Kennemer MI, Guan Y, Lee W, Carnevali P, Stinson J, Johnson S, Diao J, Yeung S, Jubb A, Ye W, Wu TD, Kapadia SB, de Sauvage FJ, Gentleman RC, Stern HM, Seshagiri S, Pant KP, Modrusan Z, Ballinger DG, Zhang Z. The effects of hepatitis B virus integration into the genomes of hepatocellular carcinoma patients. Genome Res. 2012;22:593–601. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
