

Complement Component C4 Regulates the Development of Experimental Autoimmune Uveitis through a T Cell-Intrinsic Mechanism
- PMID: 28955337
- PMCID: PMC5601957
- DOI: 10.3389/fimmu.2017.01116
Complement Component C4 Regulates the Development of Experimental Autoimmune Uveitis through a T Cell-Intrinsic Mechanism
Abstract
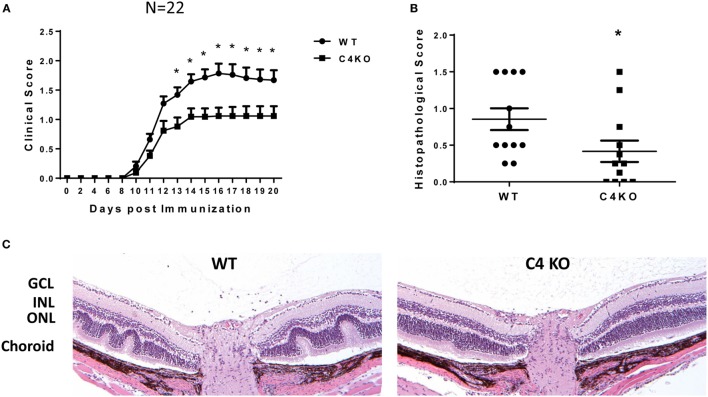
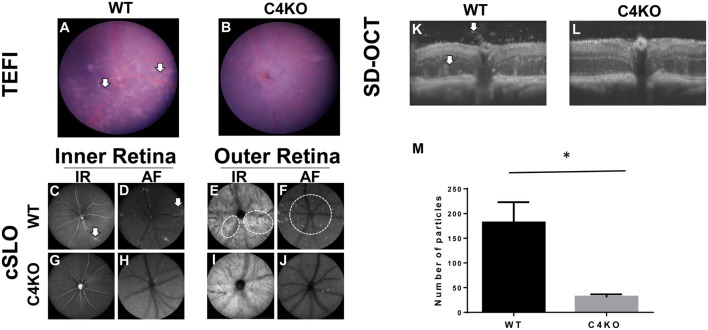
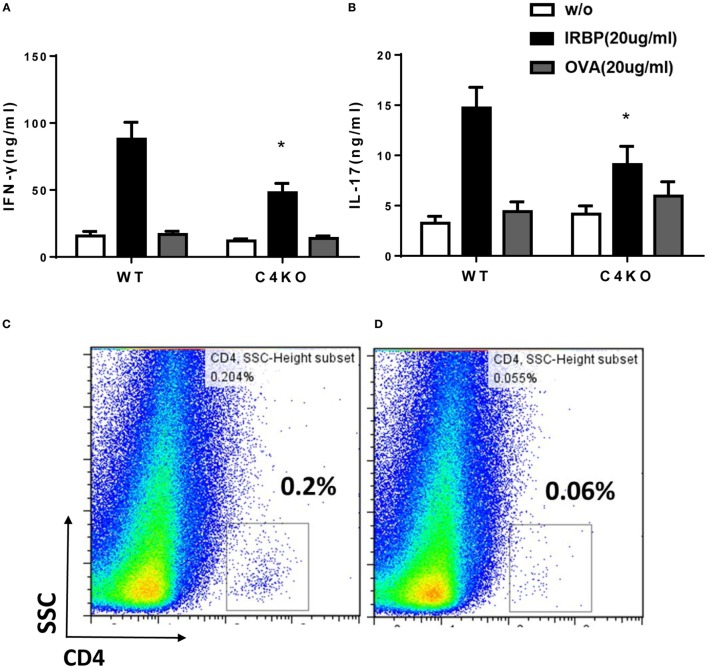
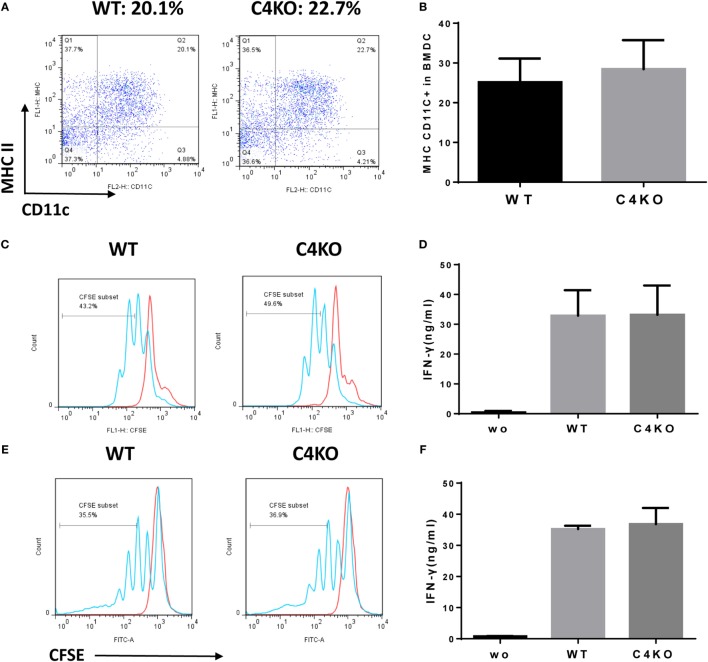
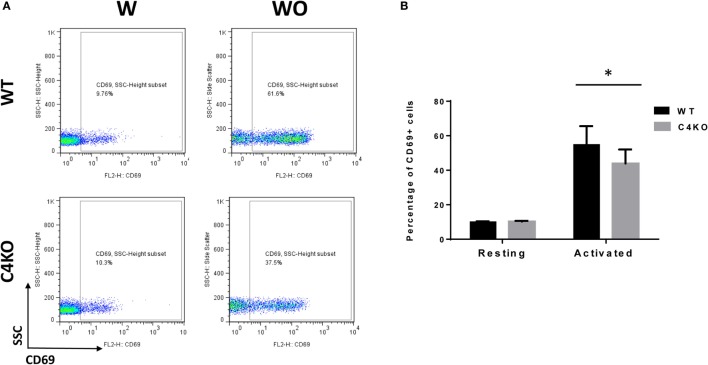
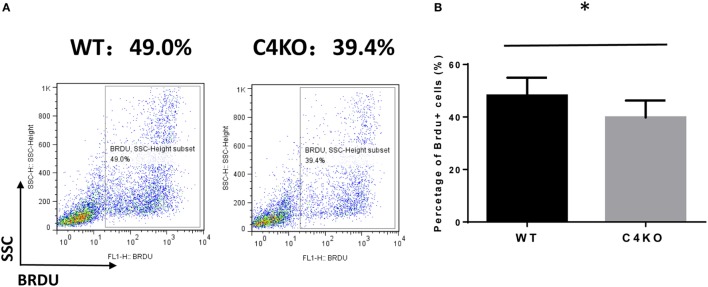
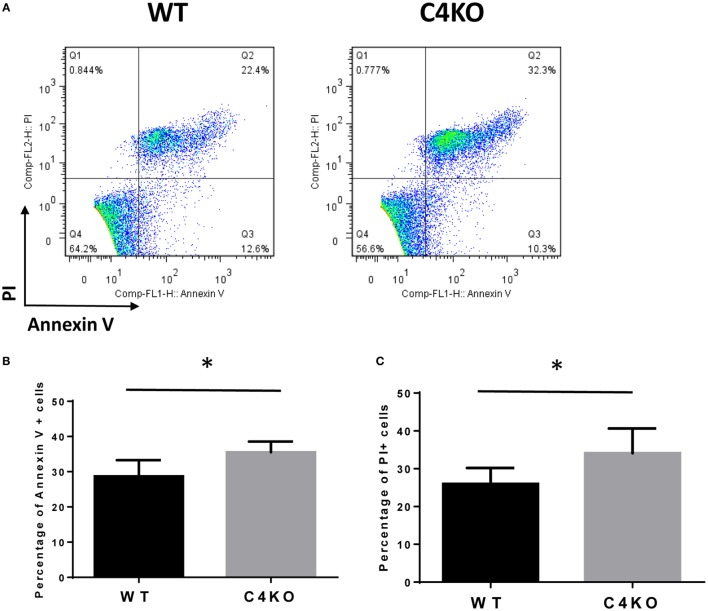
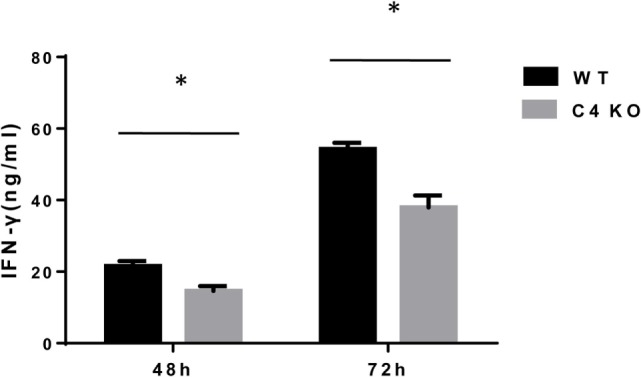
In addition to its conventional roles in the innate immune system, complement has been found to directly regulate T cells in the adaptive immune system. Complement components, including C3, C5, and factor D, are important in regulating T cell responses. However, whether complement component C4 is involved in regulating T cell responses remains unclear. In this study, we used a T cell-dependent model of autoimmunity, experimental autoimmune uveitis (EAU) to address this issue. We compared disease severity in wild-type (WT) and C4 knockout (KO) mice using indirect ophthalmoscopy, scanning laser ophthalmoscopy, spectral-domain optical coherence tomography, and histopathological analysis. We also explored the underlying mechanism by examining T cell responses in ex vivo antigen-specific recall assays and in in vitro T cell priming assays using bone marrow-derived dendritic cells, splenic dendritic cells, and T cells from WT or C4 KO mice. We found that C4 KO mice develop less severe retinal inflammation than WT mice in EAU and show reduced autoreactive T cell responses and decreased retinal T cell infiltration. We also found that T cells, but not dendritic cells, from C4 KO mice have impaired function. These results demonstrate a previously unknown role of C4 in regulating T cell responses, which affects the development of T cell-mediated autoimmunity, as exemplified by EAU. Our data could shed light on the pathogenesis of autoimmune uveitis in humans.
Keywords: C4; T cells; animal models; autoimmune uveitis; complement.
Figures
Similar articles
-
Targeting CD6 for the treatment of experimental autoimmune uveitis.J Autoimmun. 2018 Jun;90:84-93. doi: 10.1016/j.jaut.2018.02.004. Epub 2018 Feb 19. J Autoimmun. 2018. PMID: 29472120 Free PMC article.
-
Complement anaphylatoxin receptors C3aR and C5aR are required in the pathogenesis of experimental autoimmune uveitis.J Leukoc Biol. 2016 Mar;99(3):447-54. doi: 10.1189/jlb.3A0415-157R. Epub 2015 Sep 22. J Leukoc Biol. 2016. PMID: 26394814 Free PMC article.
-
Mice lacking the IFN-gamma receptor or fyn develop severe experimental autoimmune uveoretinitis characterized by different immune responses.Immunogenetics. 2005 Jun;57(5):337-43. doi: 10.1007/s00251-005-0805-3. Epub 2005 May 18. Immunogenetics. 2005. PMID: 15902435
-
Experimental autoimmune uveoretinitis (EAU)-related tissue damage and angiogenesis is reduced in CCL2⁻/⁻CX₃CR1gfp/gfp mice.Invest Ophthalmol Vis Sci. 2014 Oct 23;55(11):7572-82. doi: 10.1167/iovs.14-15495. Invest Ophthalmol Vis Sci. 2014. PMID: 25342612
-
[Intraocular inflammation and homeostasis of the eye].Nippon Ganka Gakkai Zasshi. 2009 Mar;113(3):344-77; discussion 378. Nippon Ganka Gakkai Zasshi. 2009. PMID: 19348183 Review. Japanese.
Cited by
-
Effect of bacillus subtilis strain Z15 secondary metabolites on immune function in mice.BMC Genomics. 2023 May 19;24(1):273. doi: 10.1186/s12864-023-09313-5. BMC Genomics. 2023. PMID: 37208602 Free PMC article.
-
Comparative Analysis of Porcine Follicular Fluid Proteomes of Small and Large Ovarian Follicles.Biology (Basel). 2020 May 17;9(5):101. doi: 10.3390/biology9050101. Biology (Basel). 2020. PMID: 32429601 Free PMC article.
-
Complement or insult: the emerging link between complement cascade deficiencies and pathology of myeloid malignancies.J Leukoc Biol. 2024 Nov 4;116(5):966-984. doi: 10.1093/jleuko/qiae130. J Leukoc Biol. 2024. PMID: 38836653 Review.
-
Complement 4 Aids in the Prediction of Newly Diagnosed Multiple Myeloma Outcome in Patients.Clin Med Insights Oncol. 2022 Feb 28;16:11795549221079171. doi: 10.1177/11795549221079171. eCollection 2022. Clin Med Insights Oncol. 2022. PMID: 35250324 Free PMC article.
-
Kinetic changes in microglia-related retinal transcripts in experimental autoimmune uveoretinitis (EAU) of B10.RIII mice.J Neuroinflammation. 2025 Feb 10;22(1):37. doi: 10.1186/s12974-025-03358-x. J Neuroinflammation. 2025. PMID: 39930455 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous