

Novel Role for Interleukin-17 in Enhancing Type 1 Helper T Cell Immunity in the Female Genital Tract following Mucosal Herpes Simplex Virus 2 Vaccination
- PMID: 28956763
- PMCID: PMC5686749
- DOI: 10.1128/JVI.01234-17
Novel Role for Interleukin-17 in Enhancing Type 1 Helper T Cell Immunity in the Female Genital Tract following Mucosal Herpes Simplex Virus 2 Vaccination
Abstract
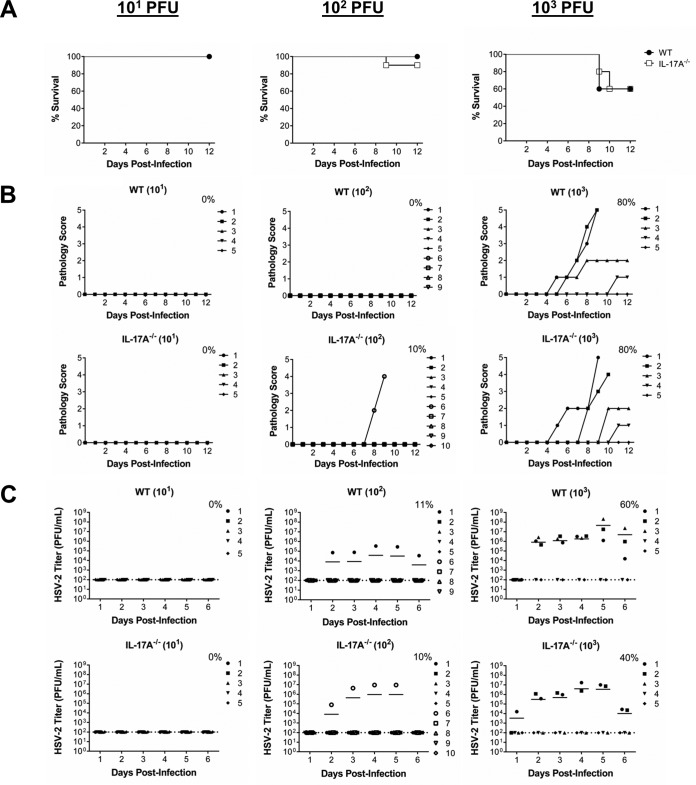
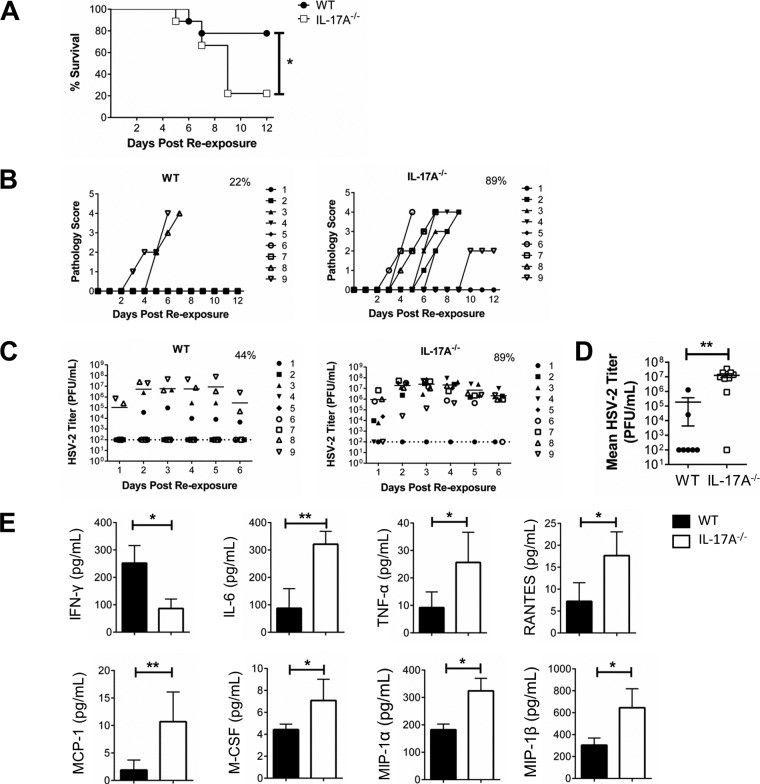
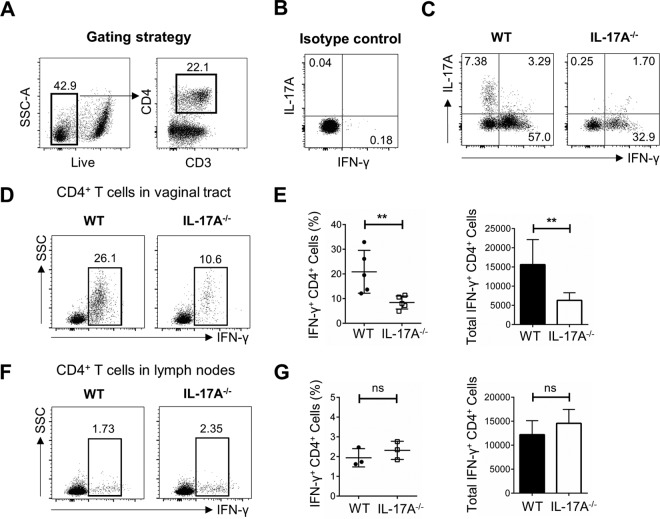
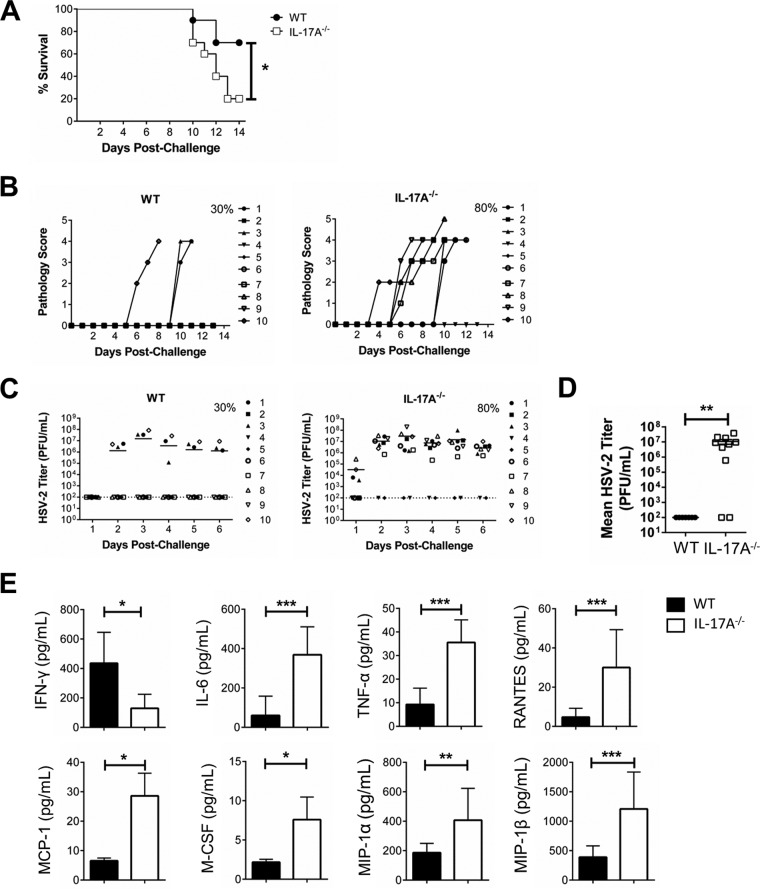
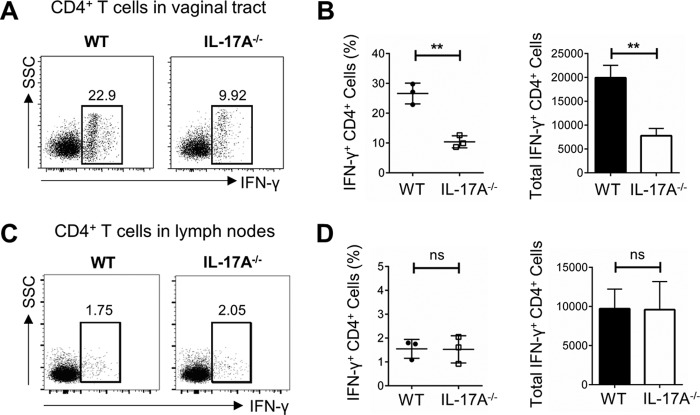
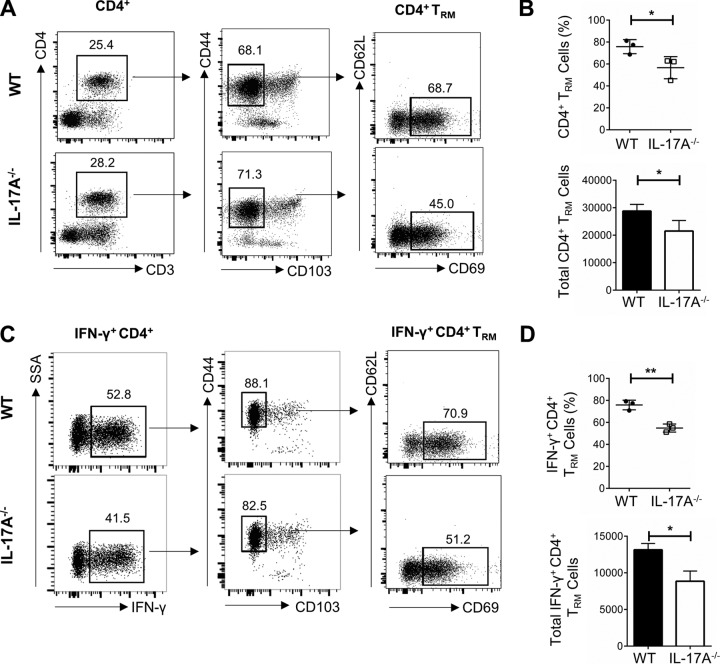
It is well established that interferon gamma (IFN-γ) production by CD4+ T cells is critical for antiviral immunity against herpes simplex virus 2 (HSV-2) genital infection. However, the role of interleukin-17A (IL-17A) production by CD4+ T cells in HSV-2 antiviral immunity is yet to be elucidated. Here we demonstrate that IL-17A plays an important role in enhancing antiviral T helper type 1 (Th1) responses in the female genital tract (FGT) and is essential for effective protection conferred by HSV-2 vaccination. While IL-17A did not play a critical role during primary genital HSV-2 infection, seen by lack of differences in susceptibility between IL-17A-deficient (IL-17A-/-) and wild-type (WT) C57BL/6 mice, it was critical for mediating antiviral responses after challenge/reexposure. Compared to WT mice, IL-17A-/- mice (i) infected intravaginally and reexposed or (ii) vaccinated intranasally and challenged intravaginally demonstrated poor outcomes. Following intravaginal HSV-2 reexposure or challenge, vaccinated IL-17A-/- mice had significantly higher mortality, greater disease severity, higher viral shedding, and higher levels of proinflammatory cytokines and chemokines in vaginal secretions. Furthermore, IL-17A-/- mice had impaired Th1 cell responses after challenge/reexposure, with significantly lower proportions of vaginal IFN-γ+ CD4+ T cells. The impaired Th1 cell responses in IL-17A-/- mice coincided with smaller populations of IFN-γ+ CD4+ tissue resident memory T (TRM) cells in the genital tract postimmunization. Taken together, these findings describe a novel role for IL-17A in regulating antiviral IFN-γ+ Th1 cell immunity in the vaginal tract. This strategy could be exploited to enhance antiviral immunity following HSV-2 vaccination.IMPORTANCE T helper type 1 (Th1) immunity, specifically interferon gamma (IFN-γ) production by CD4+ T cells, is critical for protection against genital herpesvirus (HSV-2) infection, and enhancing this response can potentially help improve disease outcomes. Our study demonstrated that interleukin-17A (IL-17A) plays an essential role in enhancing antiviral Th1 responses in the female genital tract (FGT). We found that in the absence of IL-17A, preexposed and vaccinated mice showed poor disease outcomes and were unable to overcome HSV-2 reexposure/challenge. IL-17A-deficient mice (IL-17A-/-) had smaller populations of IFN-γ+ CD4+ tissue resident memory T (TRM) cells in the genital tract postimmunization than did wild-type (WT) mice, which coincided with attenuated Th1 responses postchallenge. This has important implications for developing effective vaccines against HSV-2, as we propose that strategies inducing IL-17A in the genital tract may promote more effective Th1 cell immunity and better overall protection.
Keywords: CD4 T cell immunity; IL-17; genital tract immunity; herpes simplex virus; mucosal immunity; sexually transmitted diseases.
Copyright © 2017 American Society for Microbiology.
Figures
References
-
- Milligan GN, Bernstein DI, Bourne N. 1998. T lymphocytes are required for protection of the vaginal mucosae and sensory ganglia of immune mice against reinfection with herpes simplex virus type 2. J Immunol 160:6093–6100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
