

Members of the Rid protein family have broad imine deaminase activity and can accelerate the Pseudomonas aeruginosa D-arginine dehydrogenase (DauA) reaction in vitro
- PMID: 28957411
- PMCID: PMC5619798
- DOI: 10.1371/journal.pone.0185544
Members of the Rid protein family have broad imine deaminase activity and can accelerate the Pseudomonas aeruginosa D-arginine dehydrogenase (DauA) reaction in vitro
Abstract
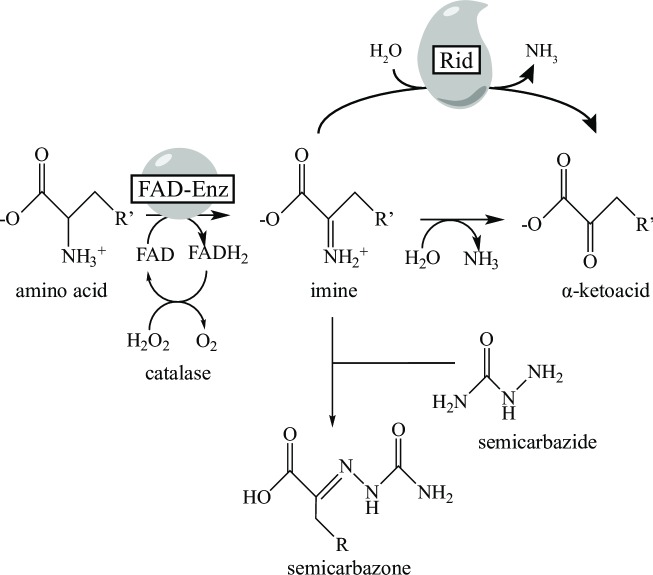
The Rid (YjgF/YER057c/UK114) protein family is a group of small, sequence diverse proteins that consists of eight subfamilies. The archetypal RidA subfamily is found in all domains, while the Rid1-7 subfamilies are present only in prokaryotes. Bacterial genomes often encode multiple members of the Rid superfamily. The best characterized member of this protein family, RidA from Salmonella enterica, is a deaminase that quenches the reactive metabolite 2-aminoacrylate generated by pyridoxal 5'-phosphate-dependent enzymes and ultimately spares certain enzymes from damage. The accumulation of 2-aminoacrylate can damage enzymes and lead to growth defects in bacteria, plants, and yeast. While all subfamily members have been annotated as imine deaminases based on the RidA characterization, experimental evidence to support this annotation exists for a single protein outside the RidA subfamily. Here we report that six proteins, spanning Rid subfamilies 1-3, deaminate a variety of imine/enamine substrates with differing specific activities. Proteins from the Rid2 and Rid3 subfamilies, but not from the RidA and Rid1 subfamilies deaminated iminoarginine, generated in situ by the Pseudomonas aeruginosa D-arginine dehydrogenase DauA. These data biochemically distinguished the subfamilies and showed Rid proteins have activity on a metabolite that is physiologically relevant in Pseudomonas and other bacteria.
Conflict of interest statement
Figures






References
-
- Burman JD, Stevenson CE, Sawers RG, Lawson DM. The crystal structure of Escherichia coli TdcF, a member of the highly conserved YjgF/YER057c/UK114 family. BMC structural biology. 2007;7:30 Epub 2007/05/18. doi: 10.1186/1472-6807-7-30 ; PubMed Central PMCID: PMCPMC1884159. - DOI - PMC - PubMed
-
- Niehaus TD, Gerdes S, Hodge-Hanson K, Zhukov A, Cooper AJ, ElBadawi-Sidhu M, et al. Genomic and experimental evidence for multiple metabolic functions in the RidA/YjgF/YER057c/UK114 (Rid) protein family. BMC genomics. 2015;16:382 Epub 2015/05/16. doi: 10.1186/s12864-015-1584-3 ; PubMed Central PMCID: PMCPMC4433059. - DOI - PMC - PubMed
-
- Browne BA, Ramos AI, Downs DM. PurF-independent phosphoribosyl amine formation in yjgF mutants of Salmonella enterica utilizes the tryptophan biosynthetic enzyme complex anthranilate synthase-phosphoribosyltransferase. Journal of bacteriology. 2006;188(19):6786–92. Epub 2006/09/19. doi: 10.1128/JB.00745-06 ; PubMed Central PMCID: PMCPMC1595518. - DOI - PMC - PubMed
-
- Christopherson MR, Schmitz GE, Downs DM. YjgF is required for isoleucine biosynthesis when Salmonella enterica is grown on pyruvate medium. J Bacteriol. 2008;190(8):3057–62. doi: 10.1128/JB.01700-07 ; PubMed Central PMCID: PMCPMC2293226. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
