

Characterization of the interactions of rabbit neonatal Fc receptor (FcRn) with rabbit and human IgG isotypes
- PMID: 28957416
- PMCID: PMC5619814
- DOI: 10.1371/journal.pone.0185662
Characterization of the interactions of rabbit neonatal Fc receptor (FcRn) with rabbit and human IgG isotypes
Abstract
Despite the increasing importance of rabbit as an animal model in pharmacological studies like investigating placental transfer of therapeutic IgGs, little is known about the molecular interaction of the rabbit neonatal Fc receptor (FcRn) with rabbit and human IgG molecules. We analyzed the interactions of the rabbit and human FcRn with rabbit and human IgG isotypes using surface plasmon resonance assay. Similar to FcRn of other species, rabbit FcRn functions in pH-dependent manner, as it binds IgGs at pH 6.0, but no binding occurs at pH 7.4. We also showed that rabbit FcRn binds rabbit IgG and human IgG1 with nearly identical affinity, whereas it has stronger interactions with the other human IgG isotypes. The similar affinity of rabbit IgG and human IgG1 for rabbit FcRn was confirmed by in vitro FcRn-mediated recycling assay. These data verify that rabbit is an appropriate animal model for analyzing the pharmacokinetics of human therapeutic monoclonal antibodies.
Conflict of interest statement
Figures

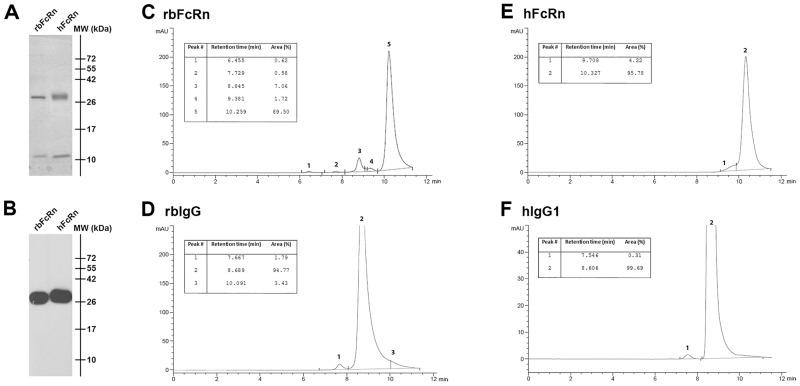
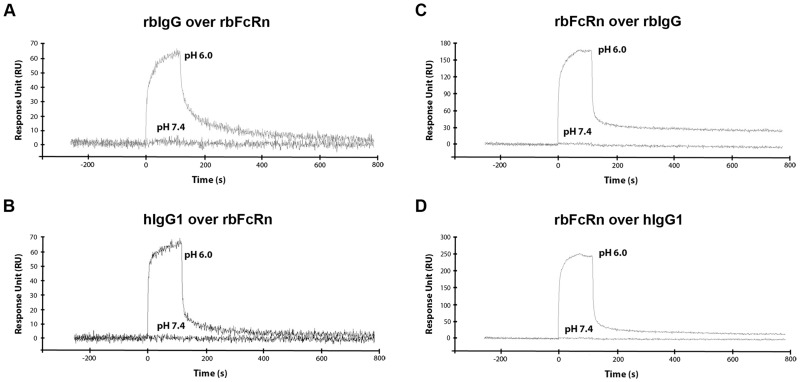
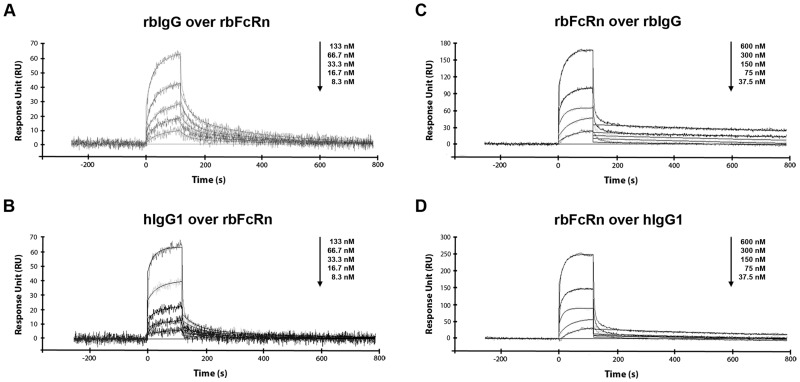
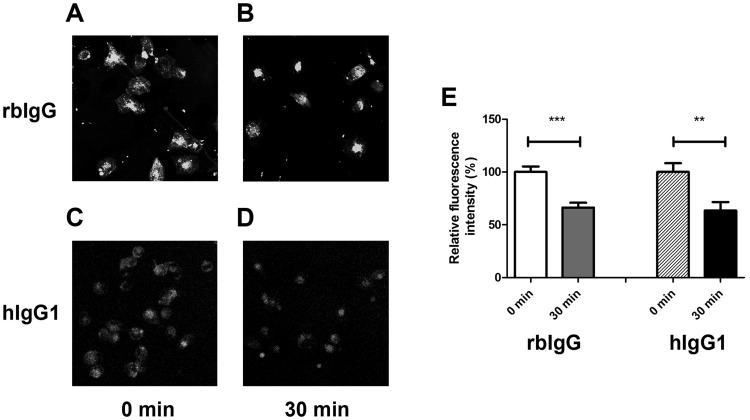
Similar articles
-
The neonatal Fc receptor (FcRn) binds independently to both sites of the IgG homodimer with identical affinity.MAbs. 2015;7(2):331-43. doi: 10.1080/19420862.2015.1008353. MAbs. 2015. PMID: 25658443 Free PMC article.
-
Crystal structure and immunoglobulin G binding properties of the human major histocompatibility complex-related Fc receptor(,).Biochemistry. 2000 Aug 15;39(32):9698-708. doi: 10.1021/bi000749m. Biochemistry. 2000. PMID: 10933786
-
Analytical FcRn affinity chromatography for functional characterization of monoclonal antibodies.MAbs. 2013 Jul-Aug;5(4):576-86. doi: 10.4161/mabs.24981. Epub 2013 May 29. MAbs. 2013. PMID: 23765230 Free PMC article.
-
Are endosomal trafficking parameters better targets for improving mAb pharmacokinetics than FcRn binding affinity?Mol Immunol. 2013 Dec;56(4):660-74. doi: 10.1016/j.molimm.2013.05.008. Epub 2013 Aug 2. Mol Immunol. 2013. PMID: 23917469 Review.
-
In Translation: FcRn across the Therapeutic Spectrum.Int J Mol Sci. 2021 Mar 17;22(6):3048. doi: 10.3390/ijms22063048. Int J Mol Sci. 2021. PMID: 33802650 Free PMC article. Review.
Cited by
-
Development of a physiologically-based pharmacokinetic model for ocular disposition of monoclonal antibodies in rabbits.J Pharmacokinet Pharmacodyn. 2020 Dec;47(6):597-612. doi: 10.1007/s10928-020-09713-0. Epub 2020 Sep 2. J Pharmacokinet Pharmacodyn. 2020. PMID: 32876799 Free PMC article.
-
Allogenic Fc Domain-Facilitated Uptake of IgG in Nasal Lamina Propria: Friend or Foe for Intranasal CNS Delivery?Pharmaceutics. 2018 Jul 26;10(3):107. doi: 10.3390/pharmaceutics10030107. Pharmaceutics. 2018. PMID: 30050027 Free PMC article.
-
Structural and Functional Analysis of CEX Fractions Collected from a Novel Avastin® Biosimilar Candidate and Its Innovator: A Comparative Study.Pharmaceutics. 2022 Jul 28;14(8):1571. doi: 10.3390/pharmaceutics14081571. Pharmaceutics. 2022. PMID: 36015197 Free PMC article.
-
Preventing lung pathology and mortality in rabbit Staphylococcus aureus pneumonia models with cytotoxin-neutralizing monoclonal IgGs penetrating the epithelial lining fluid.Sci Rep. 2019 Mar 29;9(1):5339. doi: 10.1038/s41598-019-41826-6. Sci Rep. 2019. PMID: 30926865 Free PMC article.
-
Physiologically based pharmacokinetic models for systemic disposition of protein therapeutics in rabbits.Front Pharmacol. 2024 Aug 28;15:1427325. doi: 10.3389/fphar.2024.1427325. eCollection 2024. Front Pharmacol. 2024. PMID: 39263566 Free PMC article.
References
-
- Simister NE, Mostov KE. An Fc receptor structurally related to MHC class I antigens. Nature. 1989;337(6203):184–7. doi: 10.1038/337184a0 . - DOI - PubMed
-
- Roopenian DC, Akilesh S. FcRn: the neonatal Fc receptor comes of age. Nat Rev Immunol. 2007;7(9):715–25. doi: 10.1038/nri2155 . - DOI - PubMed
-
- Baker K, Rath T, Pyzik M, Blumberg RS. The Role of FcRn in Antigen Presentation. Frontiers in immunology. 2014;5:408 doi: 10.3389/fimmu.2014.00408 . - DOI - PMC - PubMed
-
- Schneider Z, Jani PK, Szikora B, Vegh A, Kovesdi D, Ilias A, et al. Overexpression of Bovine FcRn in Mice Enhances T-Dependent Immune Responses by Amplifying T Helper Cell Frequency and Germinal Center Enlargement in the Spleen. Frontiers in immunology. 2015;6:357 doi: 10.3389/fimmu.2015.00357 . - DOI - PMC - PubMed
-
- Cervenak J, Kurrle R, Kacskovics I. Accelerating antibody discovery using transgenic animals overexpressing the neonatal Fc receptor as a result of augmented humoral immunity. Immunological reviews. 2015;268(1):269–87. doi: 10.1111/imr.12364 . - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
