

Isoform Evolution in Primates through Independent Combination of Alternative RNA Processing Events
- PMID: 28957512
- PMCID: PMC5850651
- DOI: 10.1093/molbev/msx212
Isoform Evolution in Primates through Independent Combination of Alternative RNA Processing Events
Abstract
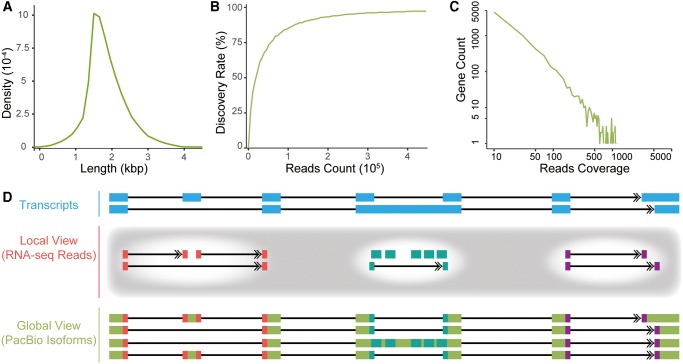
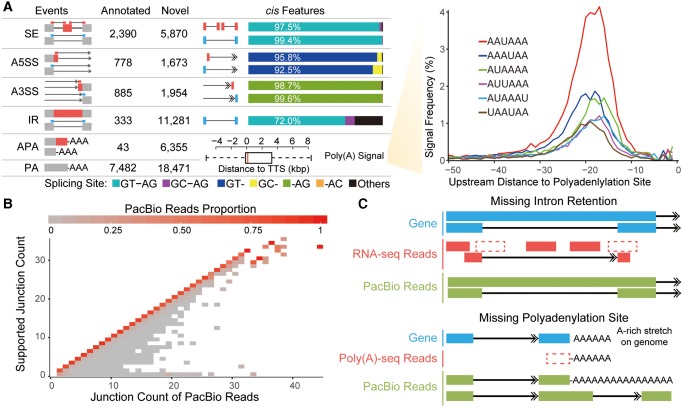
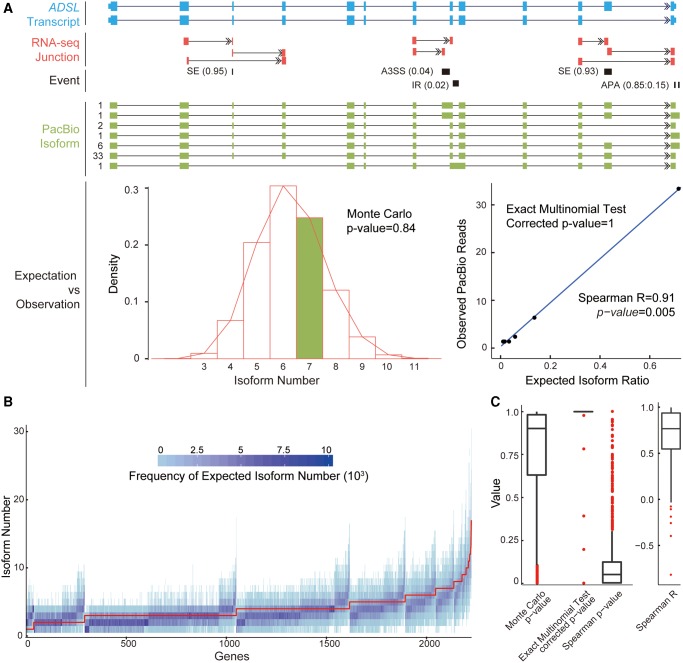
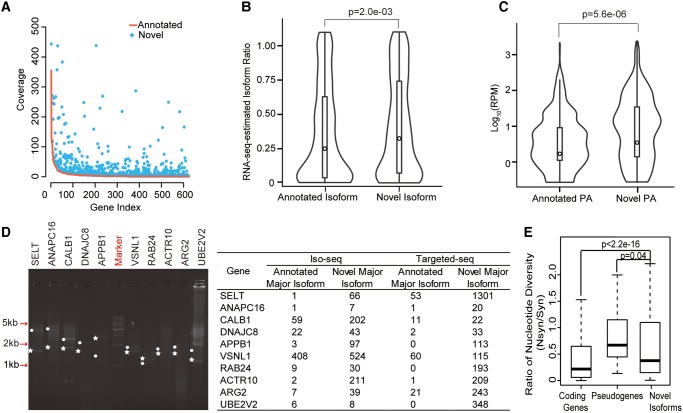
Recent RNA-seq technology revealed thousands of splicing events that are under rapid evolution in primates, whereas the reliability of these events, as well as their combination on the isoform level, have not been adequately addressed due to its limited sequencing length. Here, we performed comparative transcriptome analyses in human and rhesus macaque cerebellum using single molecule long-read sequencing (Iso-seq) and matched RNA-seq. Besides 359 million RNA-seq reads, 4,165,527 Iso-seq reads were generated with a mean length of 14,875 bp, covering 11,466 human genes, and 10,159 macaque genes. With Iso-seq data, we substantially expanded the repertoire of alternative RNA processing events in primates, and found that intron retention and alternative polyadenylation are surprisingly more prevalent in primates than previously estimated. We then investigated the combinatorial mode of these alternative events at the whole-transcript level, and found that the combination of these events is largely independent along the transcript, leading to thousands of novel isoforms missed by current annotations. Notably, these novel isoforms are selectively constrained in general, and 1,119 isoforms have even higher expression than the previously annotated major isoforms in human, indicating that the complexity of the human transcriptome is still significantly underestimated. Comparative transcriptome analysis further revealed 502 genes encoding selectively constrained, lineage-specific isoforms in human but not in rhesus macaque, linking them to some lineage-specific functions. Overall, we propose that the independent combination of alternative RNA processing events has contributed to complex isoform evolution in primates, which provides a new foundation for the study of phenotypic difference among primates.
Keywords: PacBio sequencing; alternative RNA processing event; comparative transcriptome; independent combination; isoform evolution; primate evolution.
© The Author 2017. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Barbosa-Morais NL, Irimia M, Pan Q, Xiong HY, Gueroussov S, Lee LJ, Slobodeniuc V, Kutter C, Watt S, Colak R, et al. 2012. The evolutionary landscape of alternative splicing in vertebrate species. Science 3386114: 1587–1593. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
