

Systems Approach Reveals Nuclear Factor Erythroid 2-Related Factor 2/Protein Kinase R Crosstalk in Human Cutaneous Leishmaniasis
- PMID: 28959260
- PMCID: PMC5605755
- DOI: 10.3389/fimmu.2017.01127
Systems Approach Reveals Nuclear Factor Erythroid 2-Related Factor 2/Protein Kinase R Crosstalk in Human Cutaneous Leishmaniasis
Abstract
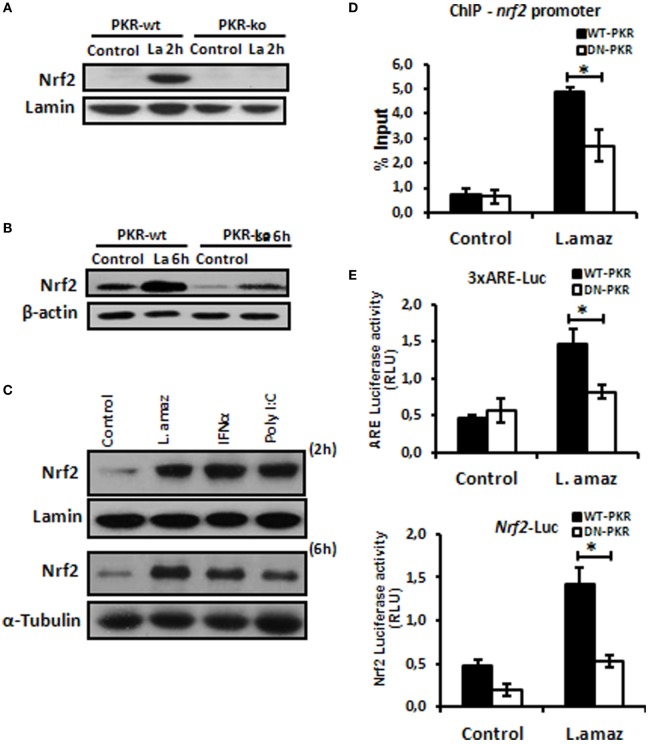
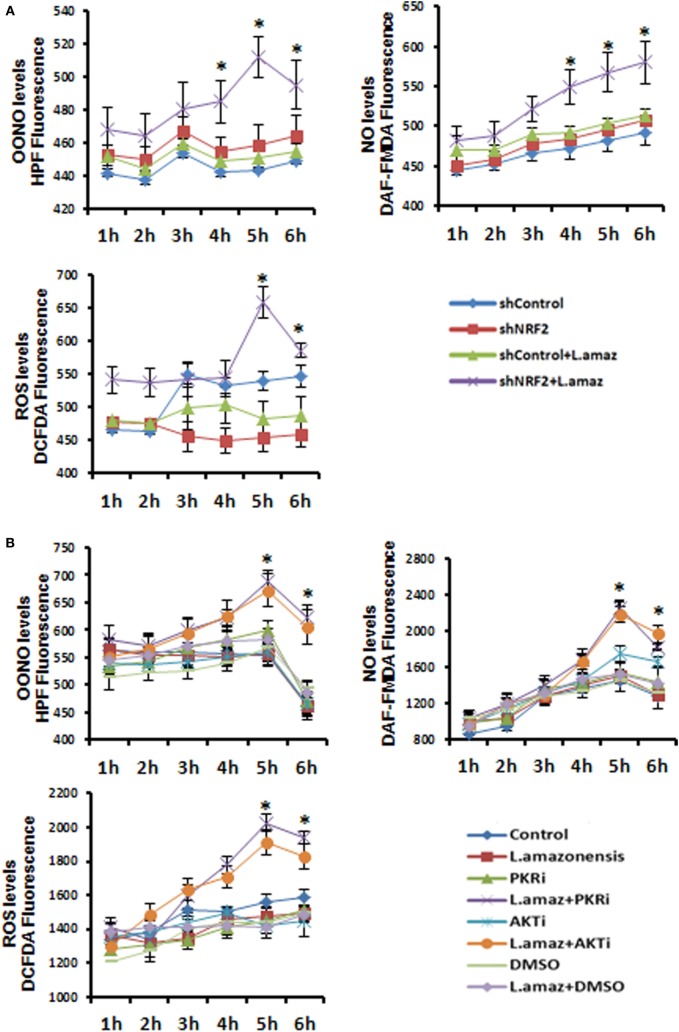
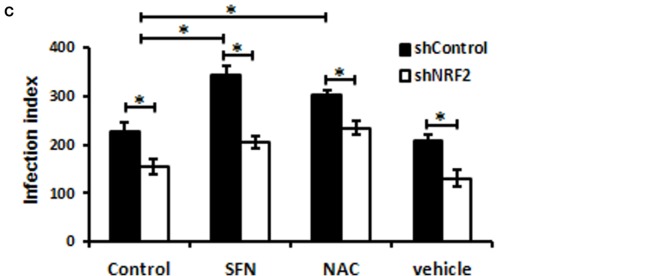
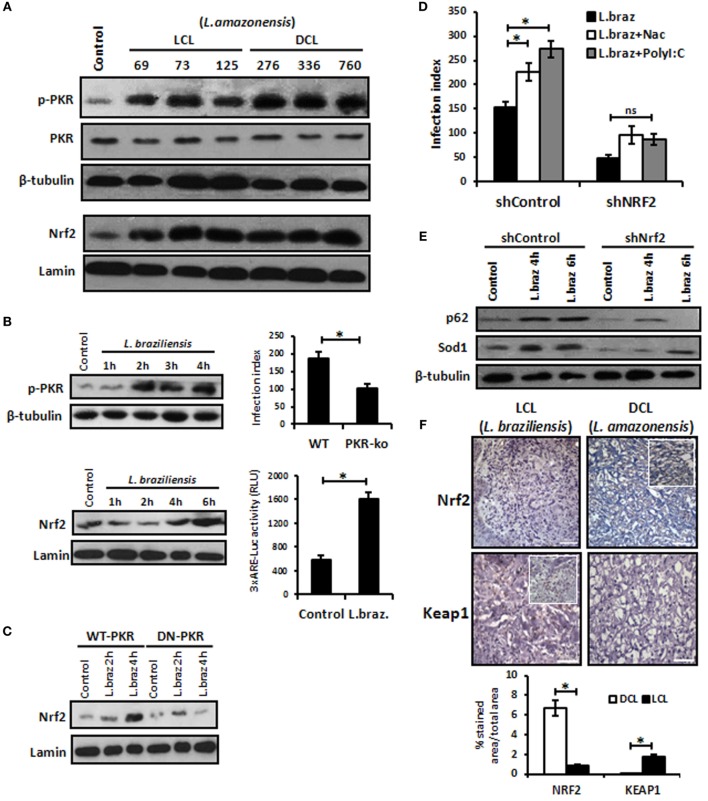
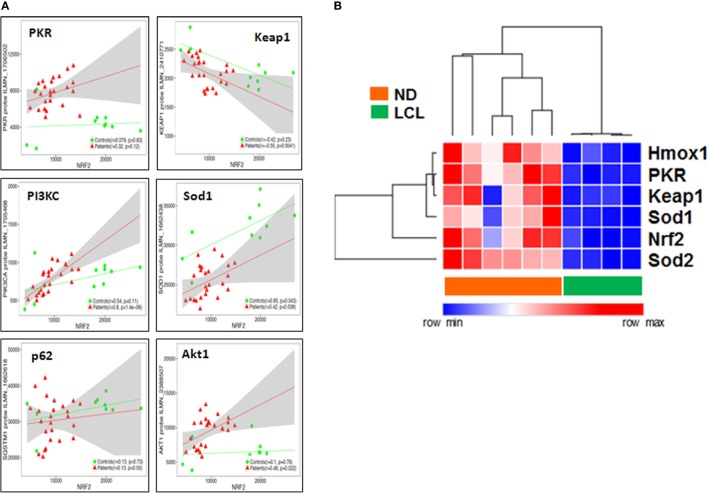
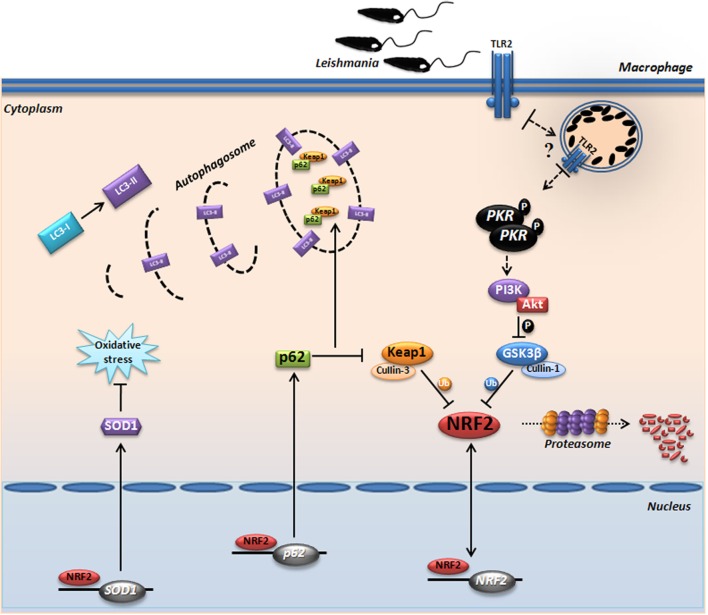
Leishmania parasites infect macrophages, causing a wide spectrum of human diseases, from cutaneous to visceral forms. In search of novel therapeutic targets, we performed comprehensive in vitro and ex vivo mapping of the signaling pathways upstream and downstream of antioxidant transcription factor [nuclear factor erythroid 2-related factor 2 (Nrf2)] in cutaneous leishmaniasis (CL), by combining functional assays in human and murine macrophages with a systems biology analysis of in situ (skin biopsies) CL patient samples. First, we show the PKR pathway controls the expression and activation of Nrf2 in Leishmania amazonensis infection in vitro. Nrf2 activation also required PI3K/Akt signaling and autophagy mechanisms. Nrf2- or PKR/Akt-deficient macrophages exhibited increased levels of ROS/RNS and reduced expression of Sod1 Nrf2-dependent gene and reduced parasite load. L. amazonensis counteracted the Nrf2 inhibitor Keap1 through the upregulation of p62 via PKR. This Nrf2/Keap1 observation was confirmed in situ in skin biopsies from Leishmania-infected patients. Next, we explored the ex vivo transcriptome in CL patients, as compared to healthy controls. We found the antioxidant response element/Nrf2 signaling pathway was significantly upregulated in CL, including downstream target p62. In silico enrichment analysis confirmed upstream signaling by interferon and PI3K/Akt, and validated our in vitro findings. Our integrated in vitro, ex vivo, and in silico approach establish Nrf2 as a central player in human cutaneous leishmaniasis and reveal Nrf2/PKR crosstalk and PI3K/Akt pathways as potential therapeutic targets.
Keywords: Leishmania; PKR; Sod1; macrophage; nuclear factor erythroid 2-related factor 2.
Figures



Similar articles
-
Human cutaneous leishmaniasis: interferon-dependent expression of double-stranded RNA-dependent protein kinase (PKR) via TLR2.FASEB J. 2011 Dec;25(12):4162-73. doi: 10.1096/fj.11-185165. Epub 2011 Aug 16. FASEB J. 2011. PMID: 21846836
-
Proteomic Analysis Reveals a Predominant NFE2L2 (NRF2) Signature in Canonical Pathway and Upstream Regulator Analysis of Leishmania-Infected Macrophages.Front Immunol. 2019 Jun 28;10:1362. doi: 10.3389/fimmu.2019.01362. eCollection 2019. Front Immunol. 2019. PMID: 31316499 Free PMC article.
-
IL-27 enhances Leishmania amazonensis infection via ds-RNA dependent kinase (PKR) and IL-10 signaling.Immunobiology. 2015 Apr;220(4):437-44. doi: 10.1016/j.imbio.2014.11.006. Epub 2014 Nov 15. Immunobiology. 2015. PMID: 25466588
-
Nuclear erythroid 2-related factor 2: a novel potential therapeutic target for liver fibrosis.Food Chem Toxicol. 2013 Sep;59:421-7. doi: 10.1016/j.fct.2013.06.018. Epub 2013 Jun 20. Food Chem Toxicol. 2013. PMID: 23793039 Review.
-
Advance in Nrf2 Signaling Pathway in Leishmaniasis.Biomedicines. 2024 Nov 4;12(11):2525. doi: 10.3390/biomedicines12112525. Biomedicines. 2024. PMID: 39595091 Free PMC article. Review.
Cited by
-
Translational profiling of macrophages infected with Leishmania donovani identifies mTOR- and eIF4A-sensitive immune-related transcripts.PLoS Pathog. 2020 Jun 1;16(6):e1008291. doi: 10.1371/journal.ppat.1008291. eCollection 2020 Jun. PLoS Pathog. 2020. PMID: 32479529 Free PMC article.
-
The Potential Role of Nrf2 Signaling in Leishmania Infection Outcomes.Front Cell Infect Microbiol. 2020 Jan 10;9:453. doi: 10.3389/fcimb.2019.00453. eCollection 2019. Front Cell Infect Microbiol. 2020. PMID: 31998662 Free PMC article. Review.
-
The Good and Bad of Nrf2: An Update in Cancer and New Perspectives in COVID-19.Int J Mol Sci. 2021 Jul 26;22(15):7963. doi: 10.3390/ijms22157963. Int J Mol Sci. 2021. PMID: 34360732 Free PMC article. Review.
-
The antioxidant response favors Leishmania parasites survival, limits inflammation and reprograms the host cell metabolism.PLoS Pathog. 2021 Mar 25;17(3):e1009422. doi: 10.1371/journal.ppat.1009422. eCollection 2021 Mar. PLoS Pathog. 2021. PMID: 33765083 Free PMC article.
-
Transcription Factors Interplay Orchestrates the Immune-Metabolic Response of Leishmania Infected Macrophages.Front Cell Infect Microbiol. 2021 Apr 7;11:660415. doi: 10.3389/fcimb.2021.660415. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 33898331 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous