

Trade-offs between seed and leaf size (seed-phytomer-leaf theory): functional glue linking regenerative with life history strategies … and taxonomy with ecology?
- PMID: 28961937
- PMCID: PMC5714152
- DOI: 10.1093/aob/mcx084
Trade-offs between seed and leaf size (seed-phytomer-leaf theory): functional glue linking regenerative with life history strategies … and taxonomy with ecology?
Abstract
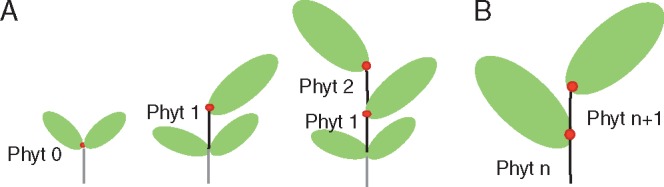
Background and aims: While the 'worldwide leaf economics spectrum' (Wright IJ, Reich PB, Westoby M, et al. 2004. The worldwide leaf economics spectrum. Nature : 821-827) defines mineral nutrient relationships in plants, no unifying functional consensus links size attributes. Here, the focus is upon leaf size, a much-studied plant trait that scales positively with habitat quality and components of plant size. The objective is to show that this wide range of relationships is explicable in terms of a seed-phytomer-leaf (SPL) theoretical model defining leaf size in terms of trade-offs involving the size, growth rate and number of the building blocks (phytomers) of which the young shoot is constructed.
Methods: Functional data for 2400+ species and English and Spanish vegetation surveys were used to explore interrelationships between leaf area, leaf width, canopy height, seed mass and leaf dry matter content (LDMC).
Key results: Leaf area was a consistent function of canopy height, LDMC and seed mass. Additionally, size traits are partially uncoupled. First, broad laminas help confer competitive exclusion while morphologically large leaves can, through dissection, be functionally small. Secondly, leaf size scales positively with plant size but many of the largest-leaved species are of medium height with basally supported leaves. Thirdly, photosynthetic stems may represent a functionally viable alternative to 'small seeds + large leaves' in disturbed, fertile habitats and 'large seeds + small leaves' in infertile ones.
Conclusions: Although key elements defining the juvenile growth phase remain unmeasured, our results broadly support SPL theory in that phytometer and leaf size are a product of the size of the initial shoot meristem (≅ seed mass) and the duration and quality of juvenile growth. These allometrically constrained traits combine to confer ecological specialization on individual species. Equally, they appear conservatively expressed within major taxa. Thus, 'evolutionary canalization' sensu Stebbins (Stebbins GL. 1974. Flowering plants: evolution above the species level . Cambridge, MA: Belknap Press) is perhaps associated with both seed and leaf development, and major taxa appear routinely specialized with respect to ecologically important size-related traits.
Keywords: Allometry; canopy height; canopy structure; evolutionary canalization; functional traits; leaf dry matter content; leaf width; photosynthetic stems; phylogeny; phytomer; seed–phytomer–leaf (SPL) theory; trade-offs.
© The Author 2017. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com
Figures






Similar articles
-
Seed size, number and strategies in annual plants: a comparative functional analysis and synthesis.Ann Bot. 2020 Nov 24;126(7):1109-1128. doi: 10.1093/aob/mcaa151. Ann Bot. 2020. PMID: 32812638 Free PMC article.
-
Combined use of leaf size and economics traits allows direct comparison of hydrophyte and terrestrial herbaceous adaptive strategies.Ann Bot. 2012 Apr;109(5):1047-53. doi: 10.1093/aob/mcs021. Epub 2012 Feb 14. Ann Bot. 2012. PMID: 22337079 Free PMC article.
-
Canopy leaf area index at its higher end: dissection of structural controls from leaf to canopy scales in bryophytes.New Phytol. 2019 Jul;223(1):118-133. doi: 10.1111/nph.15767. Epub 2019 Apr 5. New Phytol. 2019. PMID: 30821841
-
Seed size predicts global effects of small mammal seed predation on plant recruitment.Ecol Lett. 2020 Jun;23(6):1024-1033. doi: 10.1111/ele.13499. Epub 2020 Apr 5. Ecol Lett. 2020. PMID: 32249475 Review.
-
Variation in leaf photosynthetic capacity within plant canopies: optimization, structural, and physiological constraints and inefficiencies.Photosynth Res. 2023 Nov;158(2):131-149. doi: 10.1007/s11120-023-01043-9. Epub 2023 Aug 24. Photosynth Res. 2023. PMID: 37615905 Review.
Cited by
-
Shoot apical meristem and plant body organization: a cross-species comparative study.Ann Bot. 2017 Nov 10;120(5):833-843. doi: 10.1093/aob/mcx116. Ann Bot. 2017. PMID: 29136411 Free PMC article.
-
Influence of Maternal Habitat on Salt Tolerance During Germination and Growth in Zygophyllum coccineum.Plants (Basel). 2020 Nov 6;9(11):1504. doi: 10.3390/plants9111504. Plants (Basel). 2020. PMID: 33172127 Free PMC article.
-
Diversification of quantitative morphological traits in wheat.Ann Bot. 2024 Apr 10;133(3):413-426. doi: 10.1093/aob/mcad202. Ann Bot. 2024. PMID: 38195097 Free PMC article.
-
Climate as a driver of adaptive variations in ecological strategies in Arabidopsis thaliana.Ann Bot. 2018 Nov 30;122(6):935-945. doi: 10.1093/aob/mcy165. Ann Bot. 2018. PMID: 30256896 Free PMC article.
-
Phylogeny more than plant height and leaf area explains variance in seed mass.Front Plant Sci. 2023 Nov 16;14:1266798. doi: 10.3389/fpls.2023.1266798. eCollection 2023. Front Plant Sci. 2023. PMID: 38034582 Free PMC article.
References
-
- Aedo C. et al. 1980. onwards. Flora Iberica: plantas vasculares de la Península Ibérica e Islas Baleares. Madrid: Consejo Superior de Investigaciones Científicas.
-
- Angiosperm Phylogeny Group. 2016. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society 181: 1–20.
-
- Barlow PW. 1994. Rhythm, periodicity and polarity as bases for morphogenesis in plants. Biological Reviews 69: 475–525.
-
- Baskin CC, Baskin JM.. 2014. Seeds. Ecology, biogeography, and evolution of dormancy and germination, 2nd edn.San Diego: Elsevier/Academic Press.
-
- de Bello F, Berg MP, Dias ATC, et al.2015. On the need for phylogenetic ‘corrections’ in functional trait-based approaches. Folia Geobotanica 50: 349–357.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
