

Synthetic microparticles conjugated with VEGF165 improve the survival of endothelial progenitor cells via microRNA-17 inhibition
- PMID: 28963481
- PMCID: PMC5622042
- DOI: 10.1038/s41467-017-00746-7
Synthetic microparticles conjugated with VEGF165 improve the survival of endothelial progenitor cells via microRNA-17 inhibition
Abstract
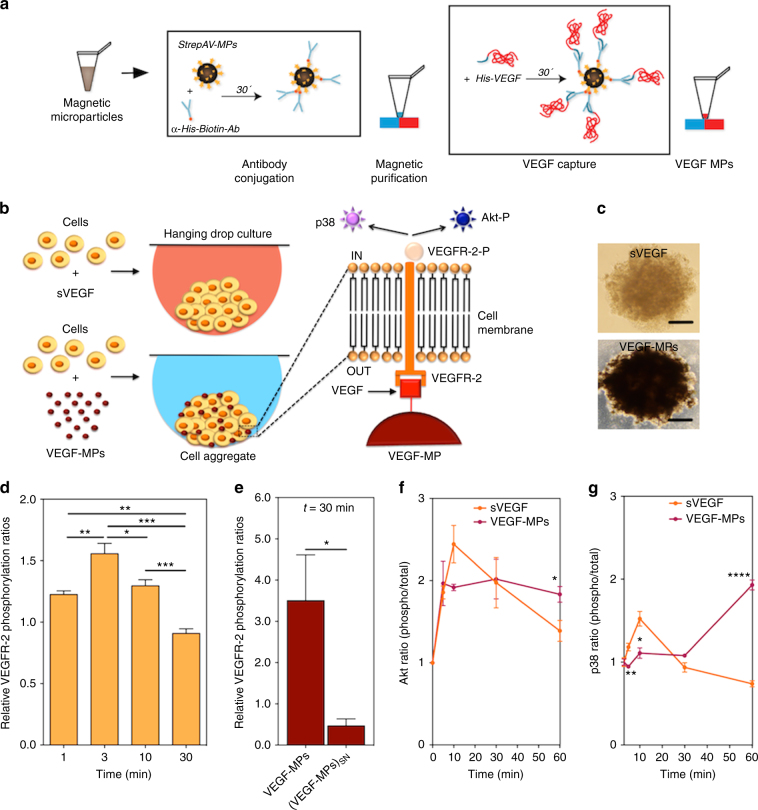
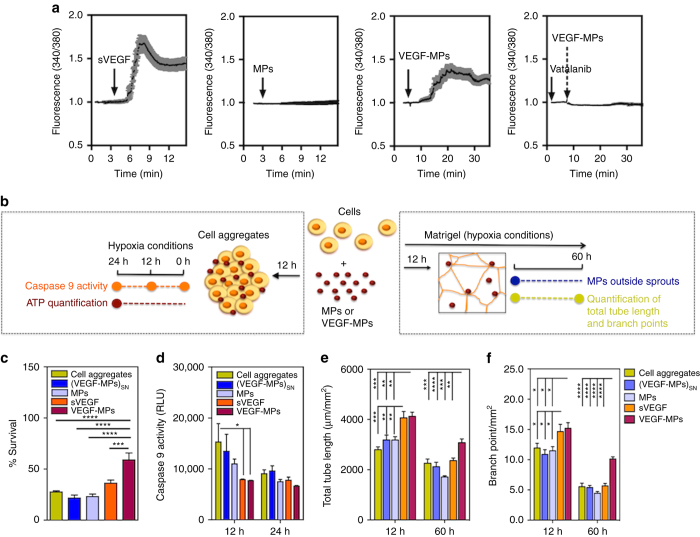
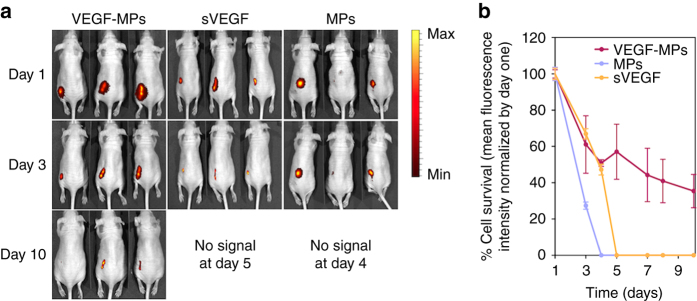
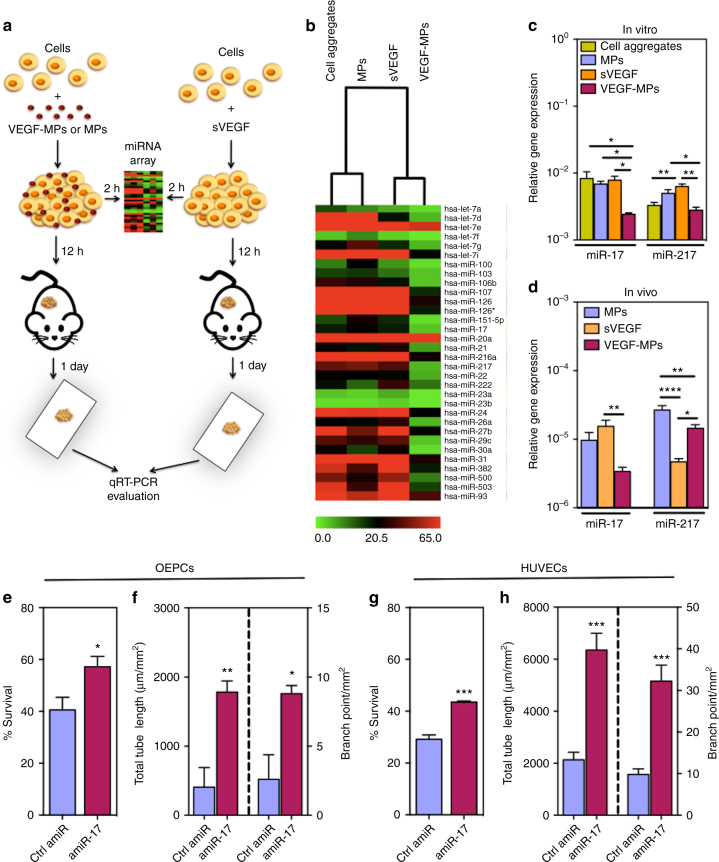
Several cell-based therapies are under pre-clinical and clinical evaluation for the treatment of ischemic diseases. Poor survival and vascular engraftment rates of transplanted cells force them to work mainly via time-limited paracrine actions. Although several approaches, including the use of soluble vascular endothelial growth factor (sVEGF)-VEGF165, have been developed in the last 10 years to enhance cell survival, they showed limited efficacy. Here, we report a pro-survival approach based on VEGF-immobilized microparticles (VEGF-MPs). VEGF-MPs prolong VEGFR-2 and Akt phosphorylation in cord blood-derived late outgrowth endothelial progenitor cells (OEPCs). In vivo, OEPC aggregates containing VEGF-MPs show higher survival than those treated with sVEGF. Additionally, VEGF-MPs decrease miR-17 expression in OEPCs, thus increasing the expression of its target genes CDKN1A and ZNF652. The therapeutic effect of OEPCs is improved in vivo by inhibiting miR-17. Overall, our data show an experimental approach to improve therapeutic efficacy of proangiogenic cells for the treatment of ischemic diseases.Soluble vascular endothelial growth factor (VEGF) enhances vascular engraftment of transplanted cells but the efficacy is low. Here, the authors show that VEGF-immobilized microparticles prolong survival of endothelial progenitors in vitro and in vivo by downregulating miR17 and upregulating CDKN1A and ZNF652.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
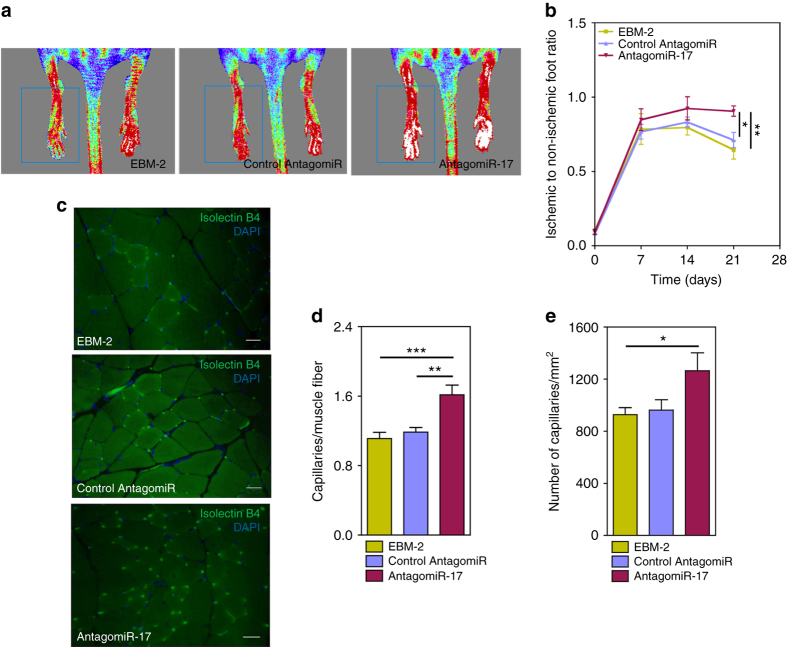
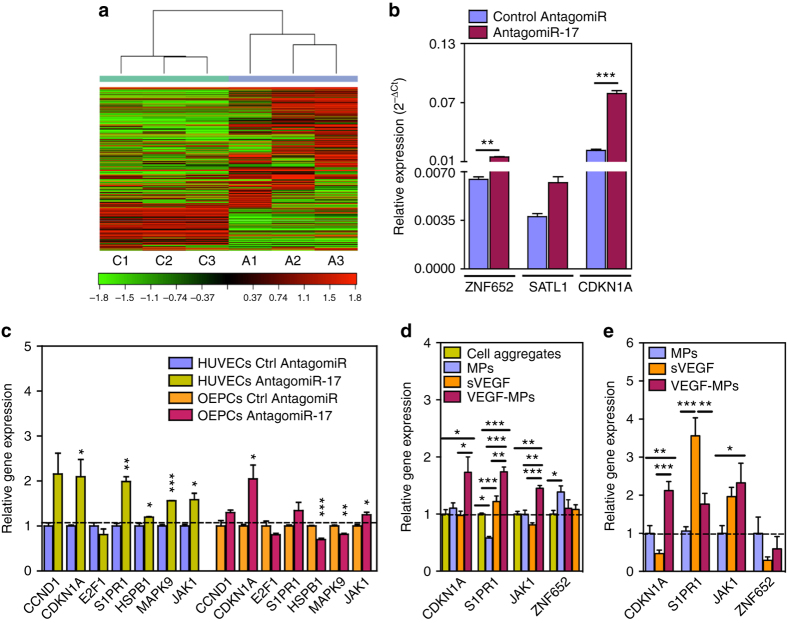
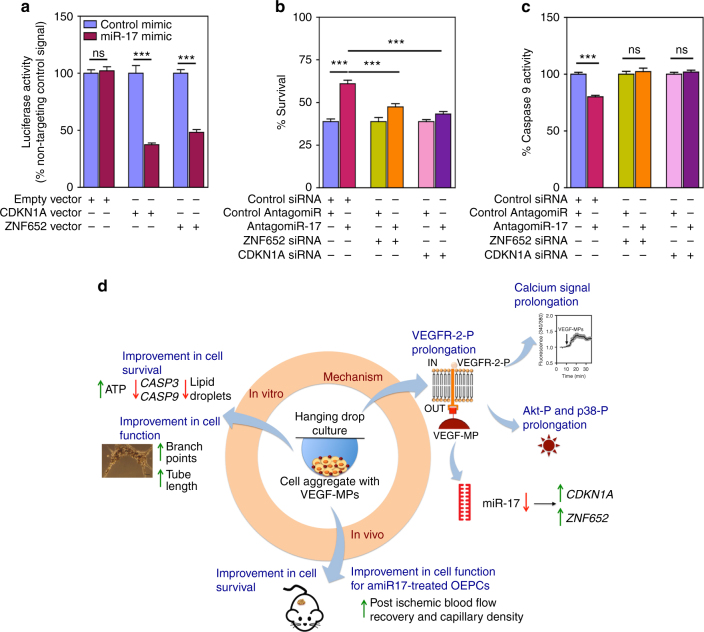
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
