

Fibroblast Growth Factor 2 as an Antifibrotic: Antagonism of Myofibroblast Differentiation and Suppression of Pro-Fibrotic Gene Expression
- PMID: 28967471
- PMCID: PMC5705586
- DOI: 10.1016/j.cytogfr.2017.09.003
Fibroblast Growth Factor 2 as an Antifibrotic: Antagonism of Myofibroblast Differentiation and Suppression of Pro-Fibrotic Gene Expression
Abstract
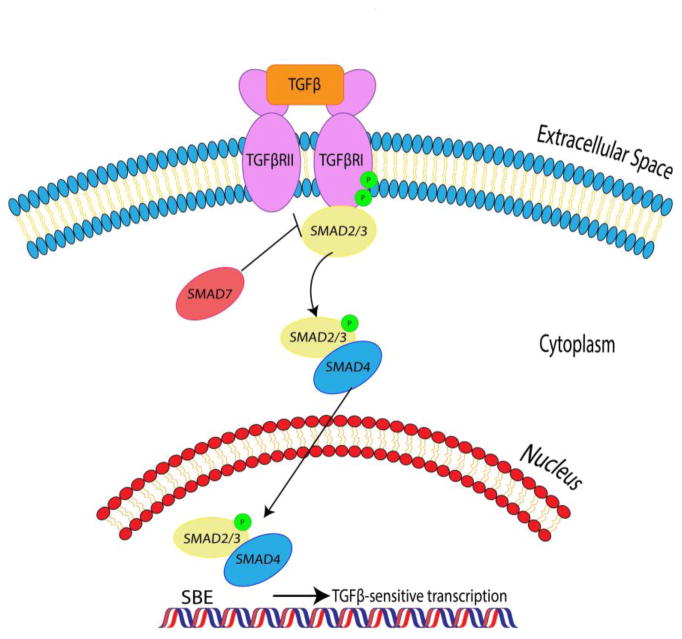
Fibrosis is a pathological condition that is characterized by the replacement of dead or damaged tissue with a nonfunctional, mechanically aberrant scar, and fibrotic pathologies account for nearly half of all deaths worldwide. The causes of fibrosis differ somewhat from tissue to tissue and pathology to pathology, but in general some of the cellular and molecular mechanisms remain constant regardless of the specific pathology in question. One of the common mechanisms underlying fibroses is the paradigm of the activated fibroblast, termed the "myofibroblast," a differentiated mesenchymal cell with demonstrated contractile activity and a high rate of collagen deposition. Fibroblast growth factor 2 (FGF2), one of the members of the mammalian fibroblast growth factor family, is a cytokine with demonstrated antifibrotic activity in non-human animal, human, and in vitro models. FGF2 is highly pleiotropic and its receptors are present on many different cell types throughout the body, lending a great deal of variety to the potential mechanisms of FGF2 effects on fibrosis. However, recent reports demonstrate that a substantial contribution to the antifibrotic effects of FGF2 comes from the inhibitory effects of FGF2 on connective tissue fibroblasts, activated myofibroblasts, and myofibroblast progenitors. FGF2 demonstrates effects antagonistic towards fibroblast activation and towards mesenchymal transition of potential myofibroblast-forming cells, as well as promotes a gene expression paradigm more reminiscent of regenerative healing, such as that which occurs in the fetal wound healing response, than fibrotic resolution. With a better understanding of the mechanisms by which FGF2 alters the wound healing cascade and results in a shift away from scar formation and towards functional tissue regeneration, we may be able to further address the critical need of therapy for varied fibrotic pathologies across myriad tissue types.
Keywords: FGF2; Fibroblast; Fibrosis; Myofibroblast; TGF-β.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors have no competing interests to report.
The authors report that there exist no relationships with any person, organization, or funding source that would constitute a conflict of interest for the submission or publication of this manuscript.
Figures


References
-
- Hinz B. Myofibroblasts. Experimental eye research. 2016;142:56–70. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
