

NLRX1 promotes immediate IRF1-directed antiviral responses by limiting dsRNA-activated translational inhibition mediated by PKR
- PMID: 28967880
- PMCID: PMC5690873
- DOI: 10.1038/ni.3853
NLRX1 promotes immediate IRF1-directed antiviral responses by limiting dsRNA-activated translational inhibition mediated by PKR
Abstract
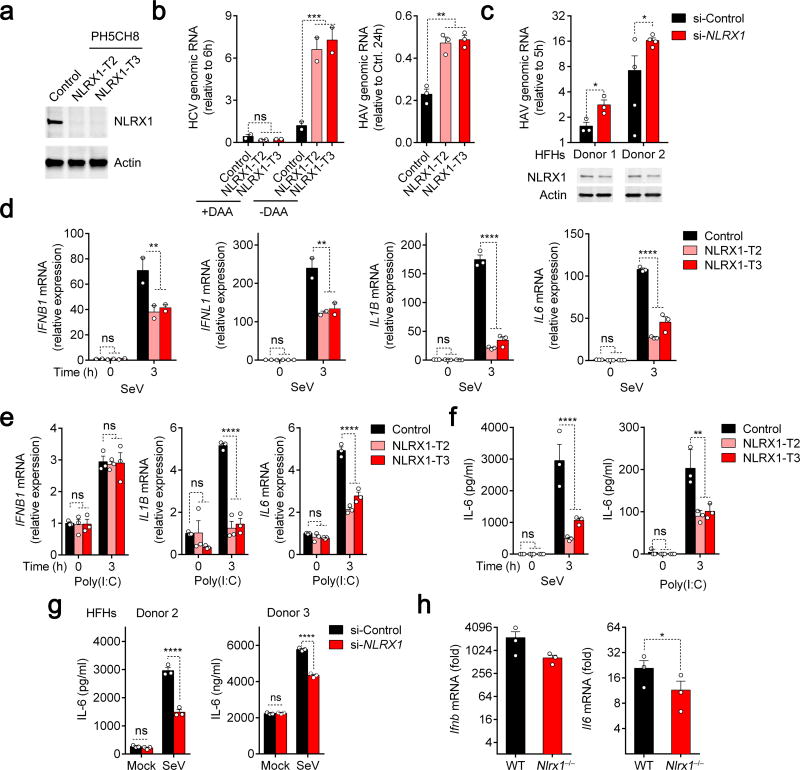
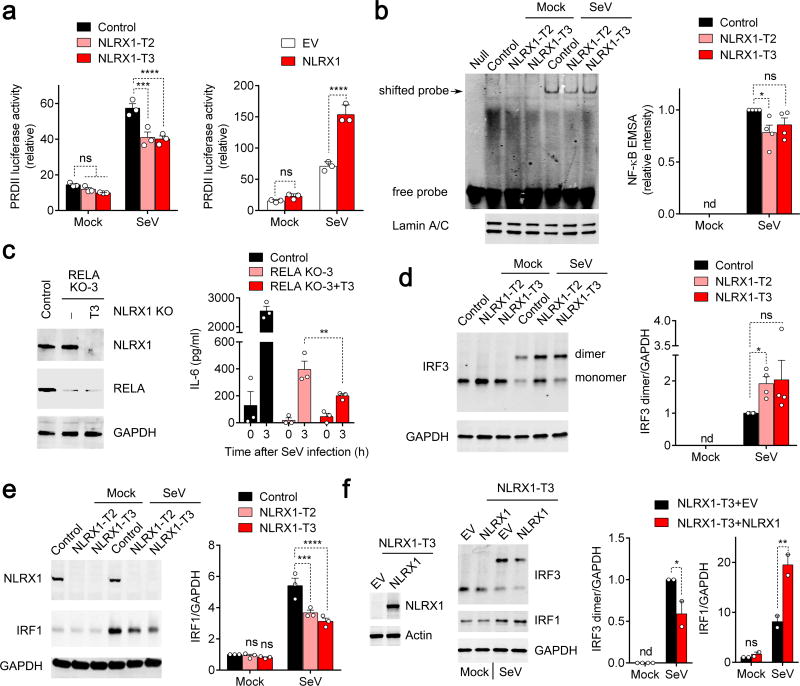
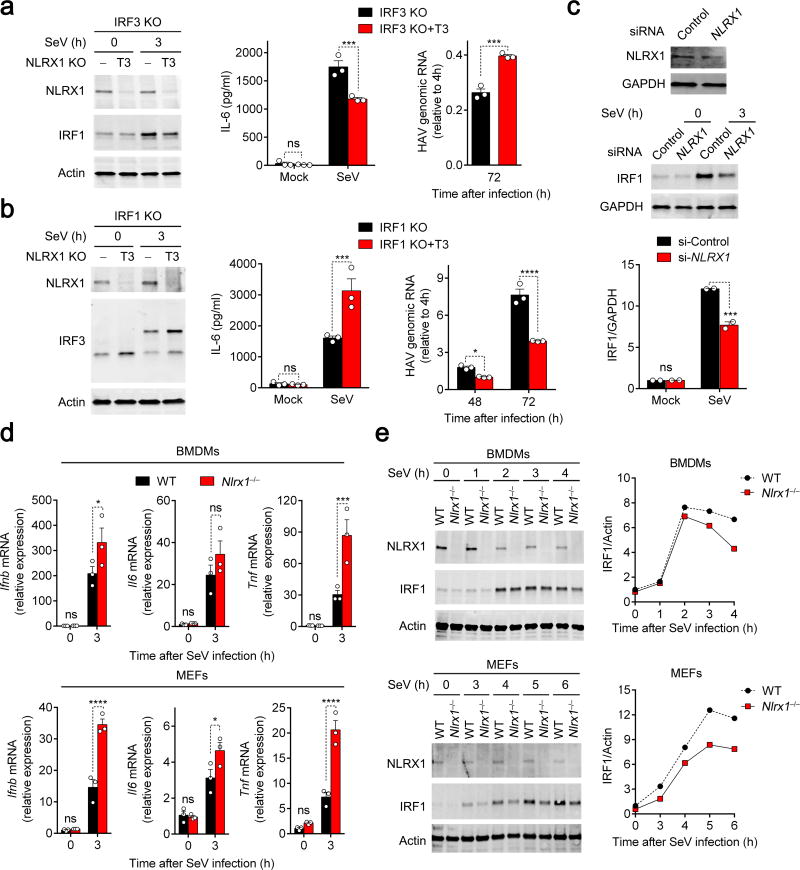
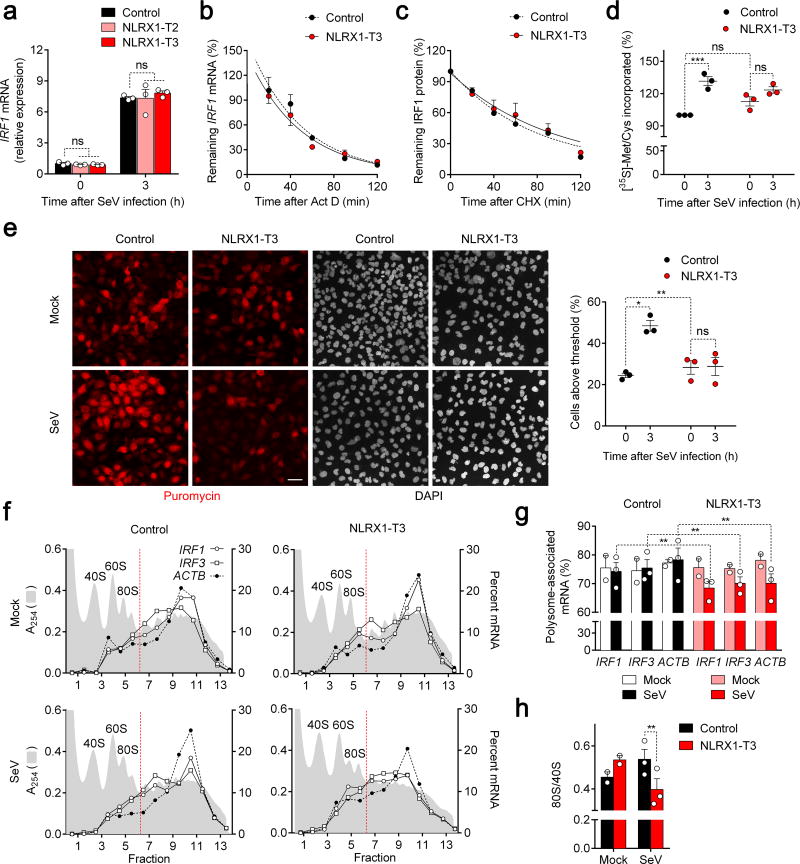
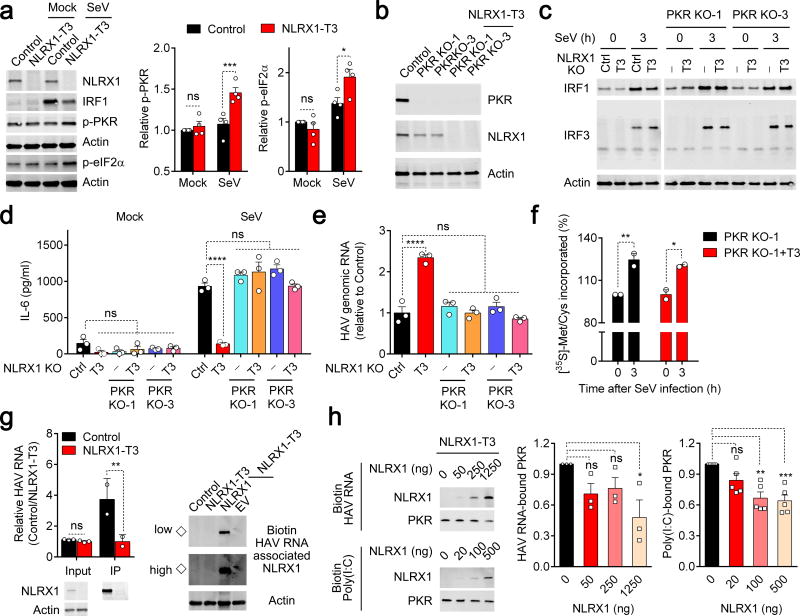
NLRX1 is unique among the nucleotide-binding-domain and leucine-rich-repeat (NLR) proteins in its mitochondrial localization and ability to negatively regulate antiviral innate immunity dependent on the adaptors MAVS and STING. However, some studies have suggested a positive regulatory role for NLRX1 in inducing antiviral responses. We found that NLRX1 exerted opposing regulatory effects on viral activation of the transcription factors IRF1 and IRF3, which might potentially explain such contradictory results. Whereas NLRX1 suppressed MAVS-mediated activation of IRF3, it conversely facilitated virus-induced increases in IRF1 expression and thereby enhanced control of viral infection. NLRX1 had a minimal effect on the transcription of IRF1 mediated by the transcription factor NF-kB and regulated the abundance of IRF1 post-transcriptionally by preventing translational shutdown mediated by the double-stranded RNA (dsRNA)-activated kinase PKR and thereby allowed virus-induced increases in the abundance of IRF1 protein.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Benko S, Magalhaes JG, Philpott DJ, Girardin SE. NLRC5 limits the activation of inflammatory pathways. J Immunol. 2010;185:1681–1691. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
