

Neuronal Expression of Opioid Gene is Controlled by Dual Epigenetic and Transcriptional Mechanism in Human Brain
- PMID: 28968778
- PMCID: PMC6887740
- DOI: 10.1093/cercor/bhx181
Neuronal Expression of Opioid Gene is Controlled by Dual Epigenetic and Transcriptional Mechanism in Human Brain
Abstract
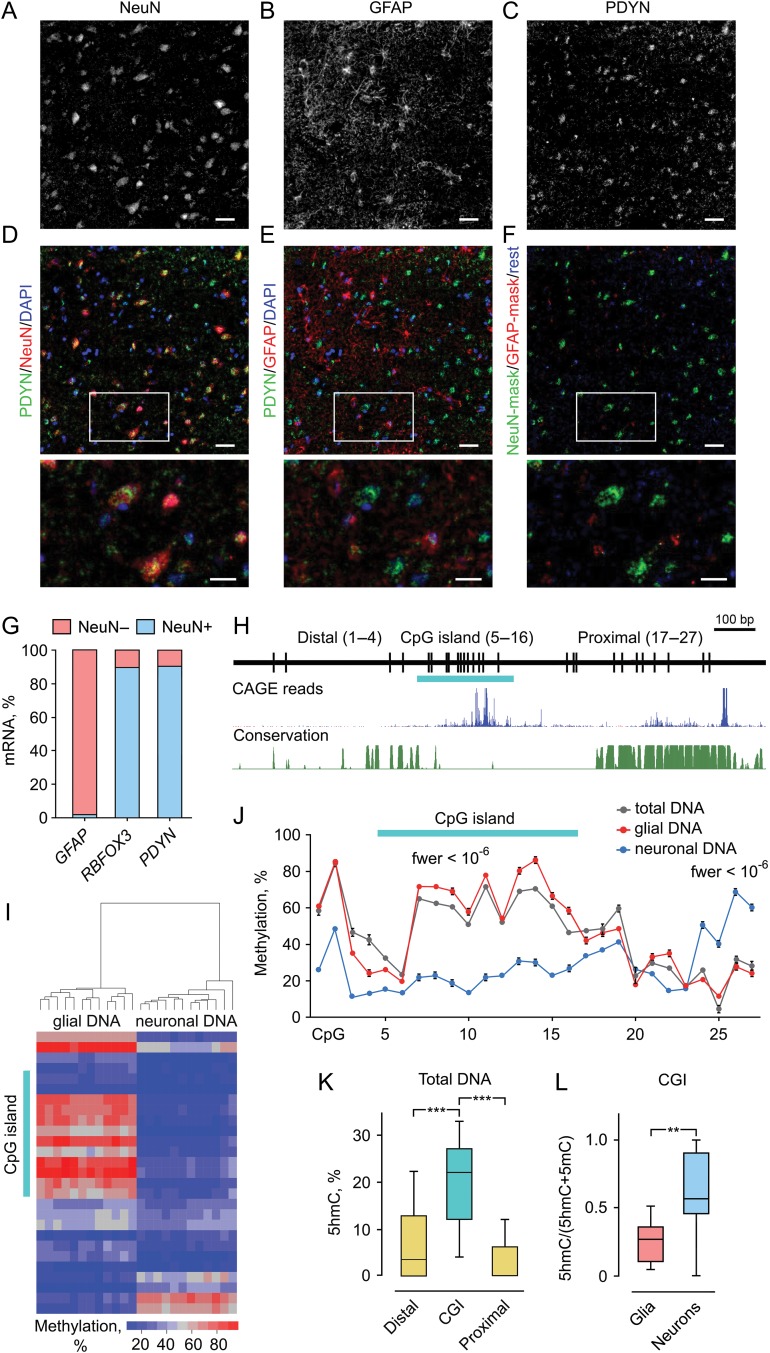
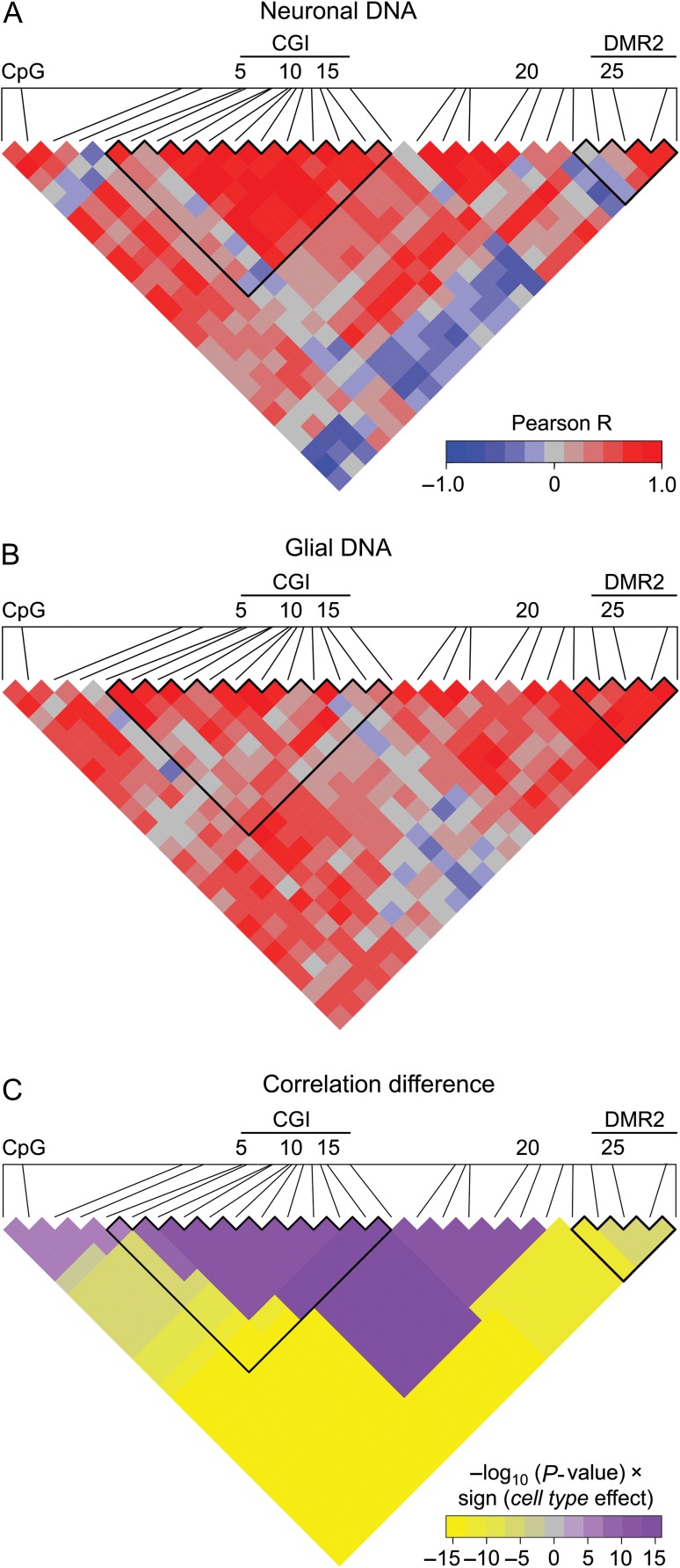
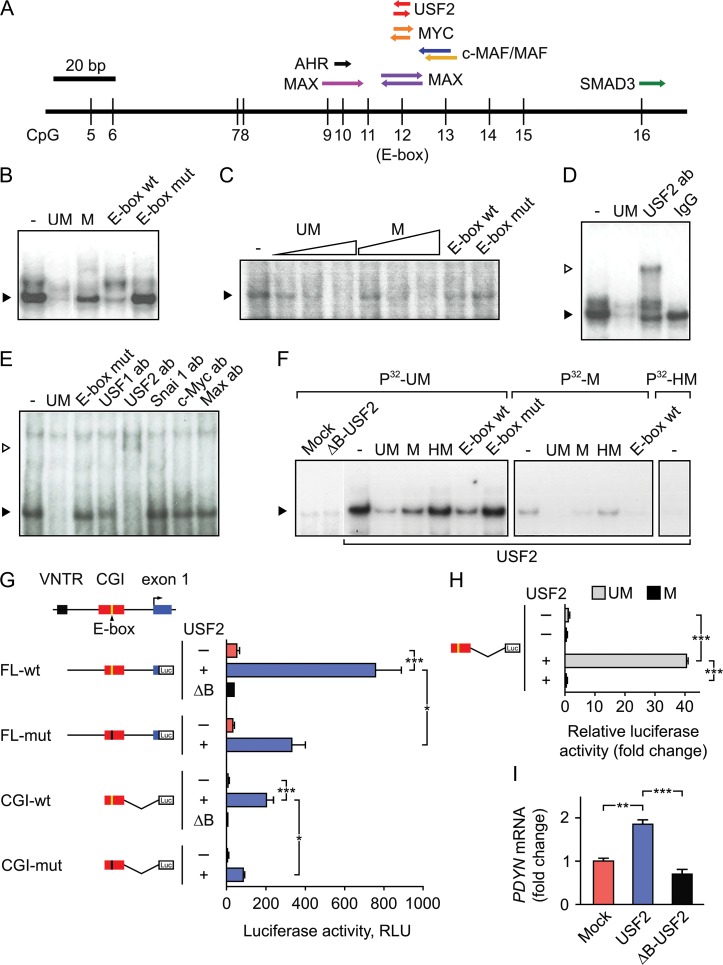
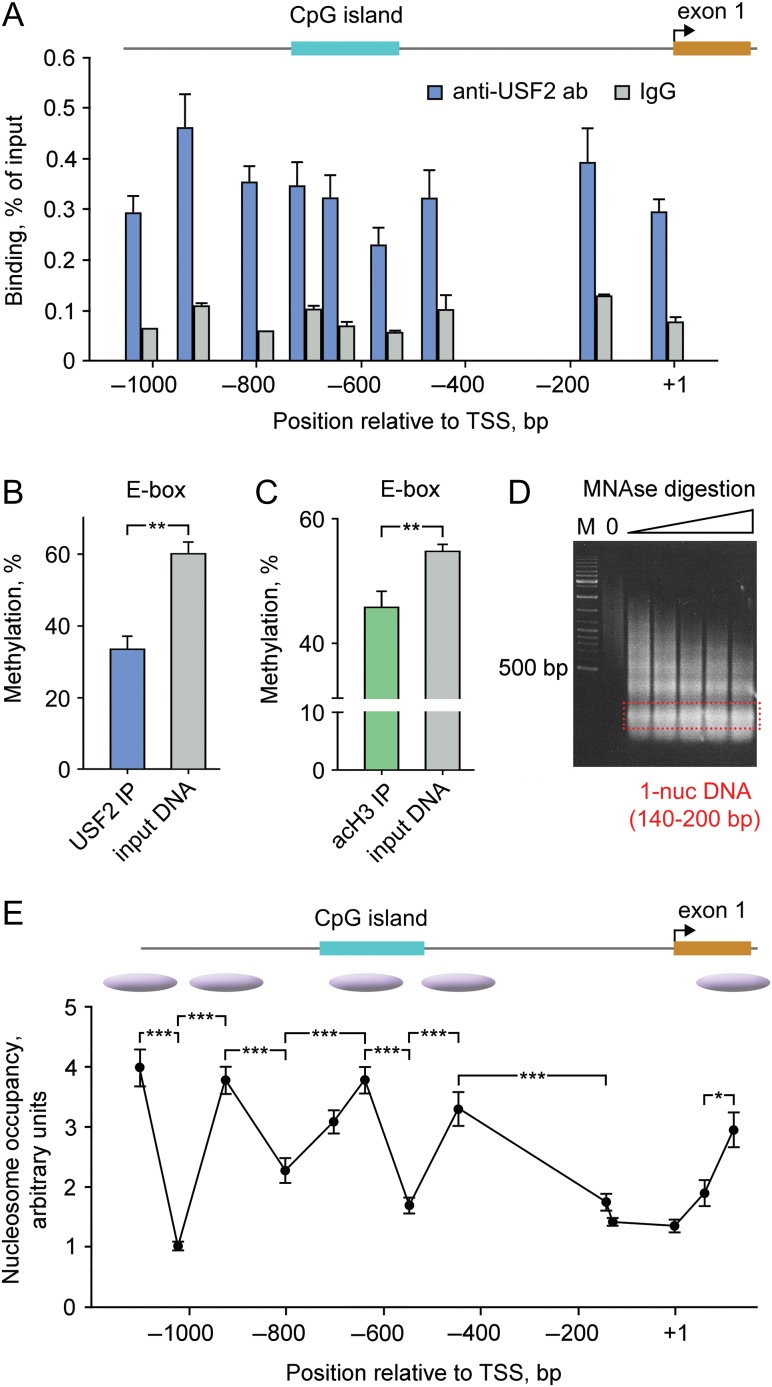
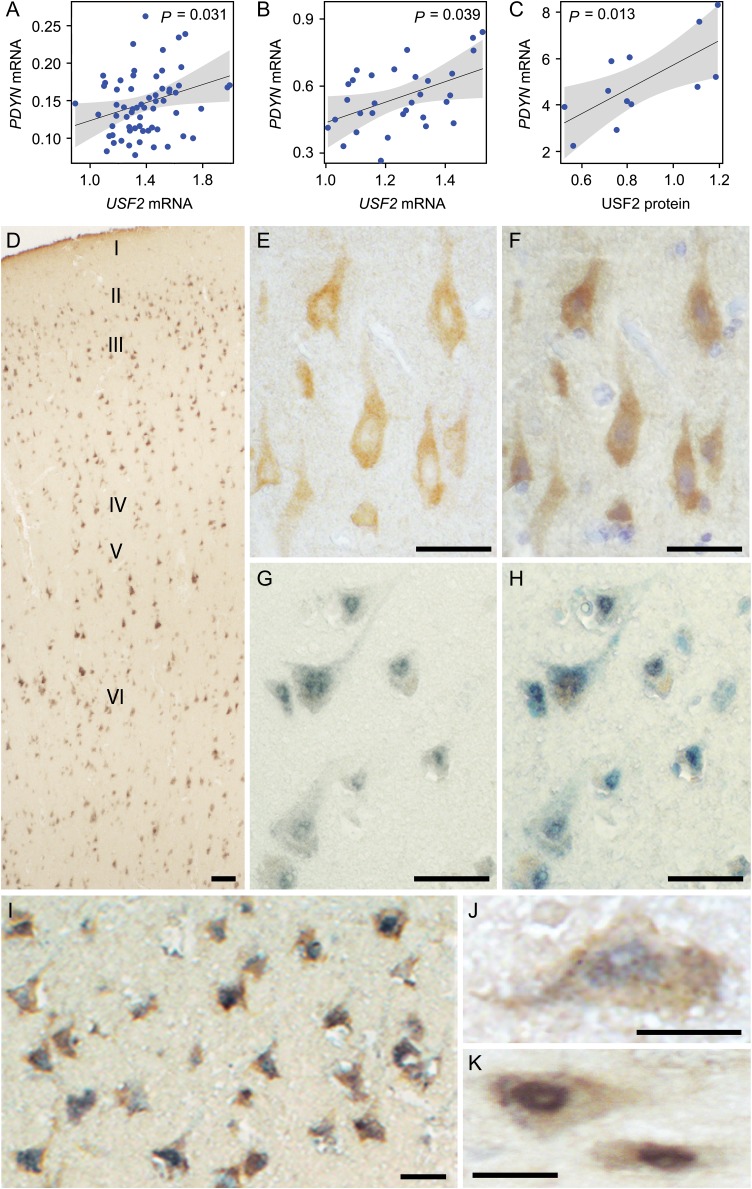
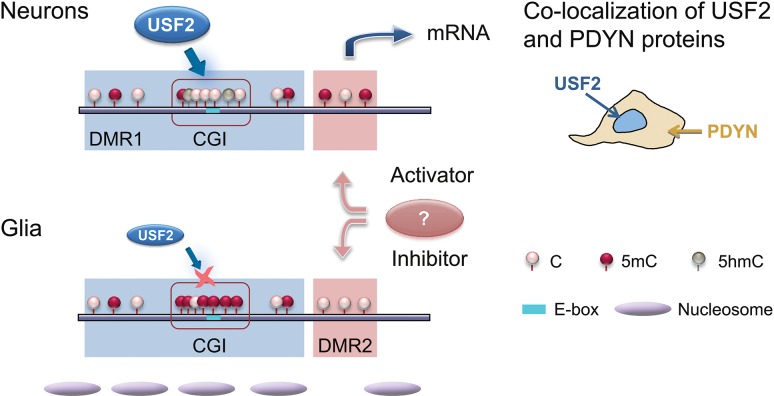
Molecular mechanisms that define patterns of neuropeptide expression are essential for the formation and rewiring of neural circuits. The prodynorphin gene (PDYN) gives rise to dynorphin opioid peptides mediating depression and substance dependence. We here demonstrated that PDYN is expressed in neurons in human dorsolateral prefrontal cortex (dlPFC), and identified neuronal differentially methylated region in PDYN locus framed by CCCTC-binding factor binding sites. A short, nucleosome size human-specific promoter CpG island (CGI), a core of this region may serve as a regulatory module, which is hypomethylated in neurons, enriched in 5-hydroxymethylcytosine, and targeted by USF2, a methylation-sensitive E-box transcription factor (TF). USF2 activates PDYN transcription in model systems, and binds to nonmethylated CGI in dlPFC. USF2 and PDYN expression is correlated, and USF2 and PDYN proteins are co-localized in dlPFC. Segregation of activatory TF and repressive CGI methylation may ensure contrasting PDYN expression in neurons and glia in human brain.
Figures
References
-
- Allis CD, Jenuwein T. 2016. The molecular hallmarks of epigenetic control. Nat Rev Genet. 17:487–500. - PubMed
-
- Babbitt CC, Silverman JS, Haygood R, Reininga JM, Rockman MV, Wray GA. 2010. Multiple functional variants in cis modulate PDYN expression. Mol Biol Evol. 27:465–479. - PubMed
-
- Bakalkin G, Watanabe H, Jezierska J, Depoorter C, Verschuuren-Bemelmans C, Bazov I, Artemenko KA, Yakovleva T, Dooijes D, Van de Warrenburg BPC, et al. 2010. Prodynorphin mutations cause the neurodegenerative disorder Spinocerebellar Ataxia Type 23 (vol 87, pg 593, 2010). Am J Human Genet. 87:736–736. - PMC - PubMed
-
- Bakalkin G, Yakovleva T, Terenius L. 1994. Prodynorphin gene expression relates to NF-kappa B factors. Brain Res Mol Brain Res. 24:301–312. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
