

Contribution of reactive oxygen species to thymineless death in Escherichia coli
- PMID: 28970486
- PMCID: PMC5705385
- DOI: 10.1038/s41564-017-0037-y
Contribution of reactive oxygen species to thymineless death in Escherichia coli
Abstract
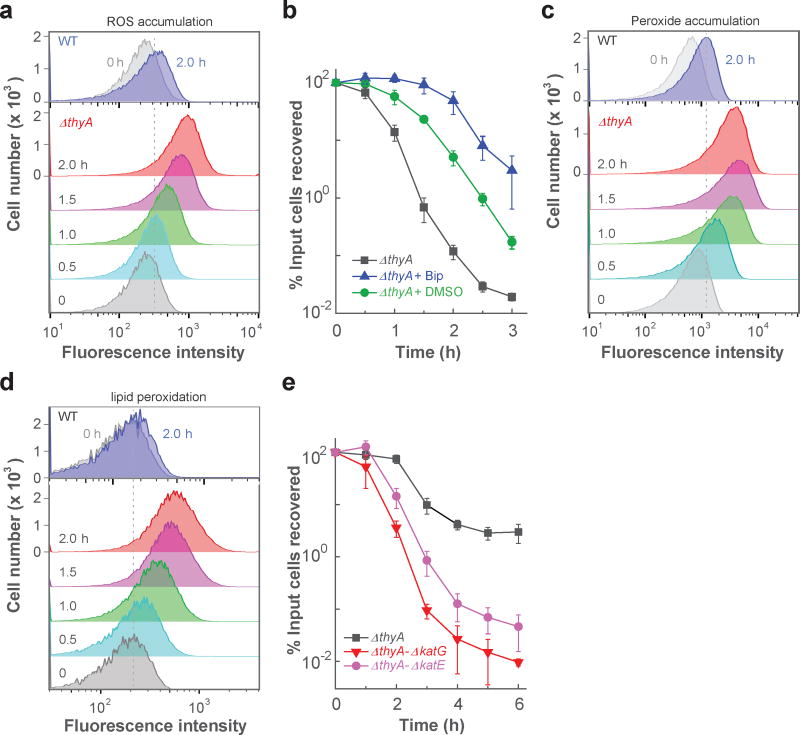
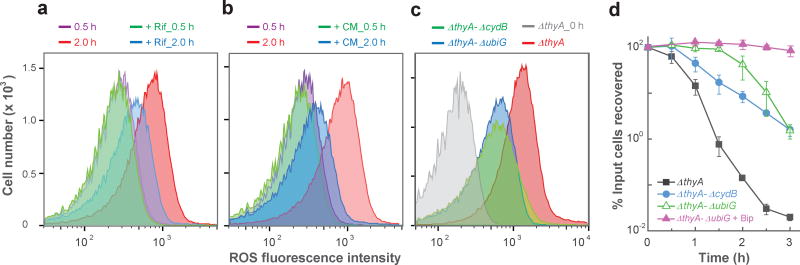
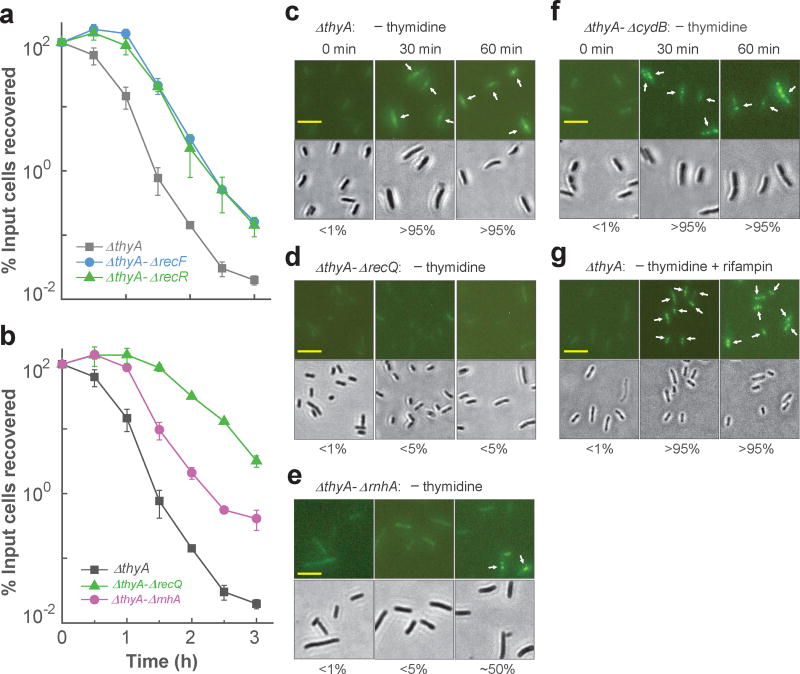
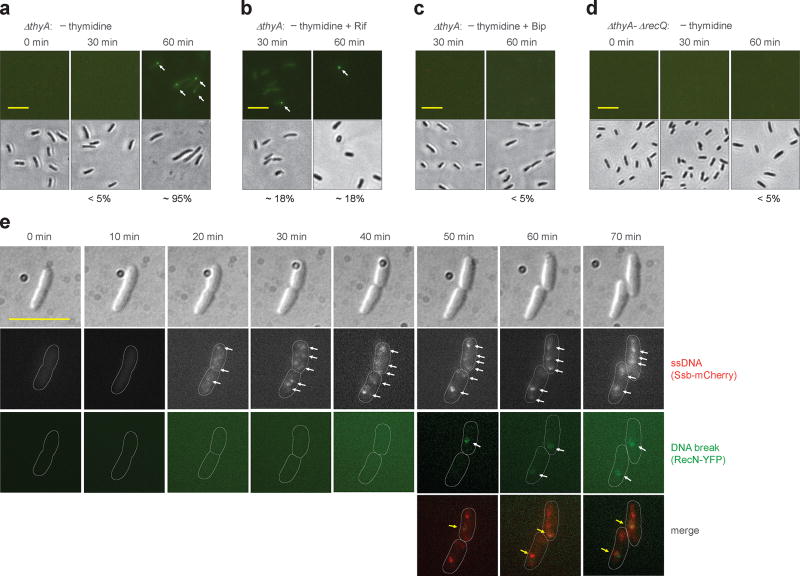
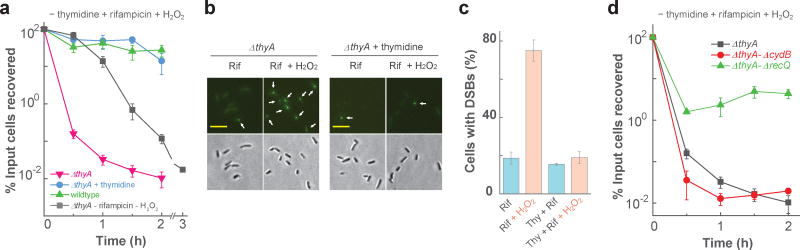
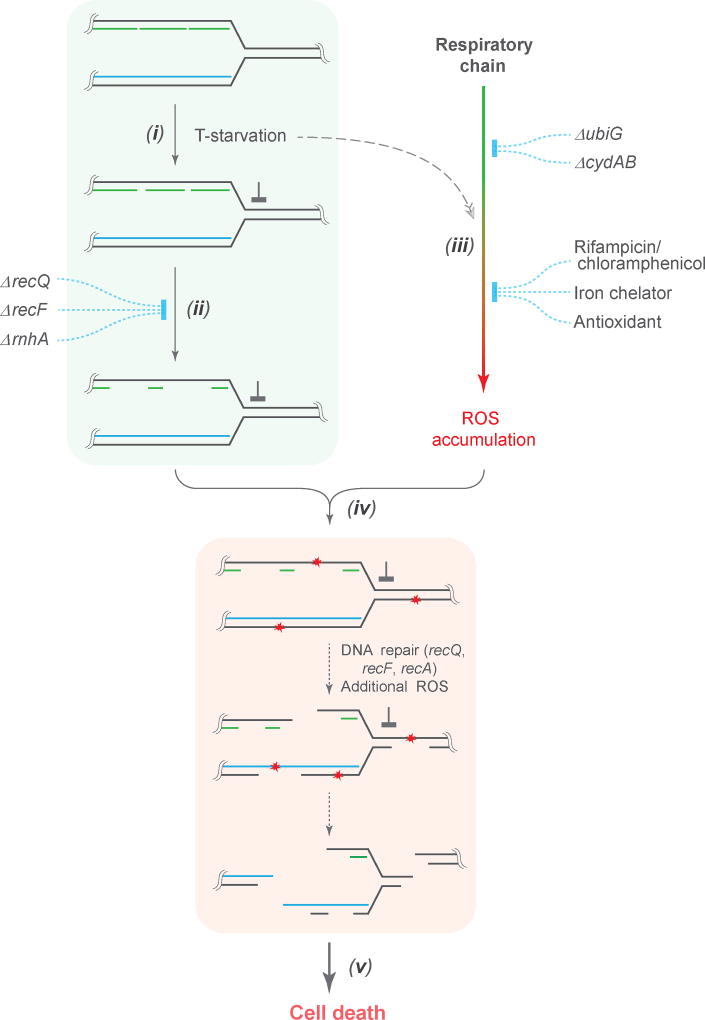
Nutrient starvation usually halts cell growth rather than causing death. Thymine starvation is exceptional, because it kills cells rapidly. This phenomenon, called thymineless death (TLD), underlies the action of several antibacterial, antimalarial, anticancer, and immunomodulatory agents. Many explanations for TLD have been advanced, with recent efforts focused on recombination proteins and replication origin (oriC) degradation. Because current proposals account for only part of TLD and because reactive oxygen species (ROS) are implicated in bacterial death due to other forms of harsh stress, we investigated the possible involvement of ROS in TLD. Here, we show that thymine starvation leads to accumulation of both single-stranded DNA regions and intracellular ROS, and interference with either event protects bacteria from double-stranded DNA breakage and TLD. Elevated levels of single-stranded DNA were necessary but insufficient for TLD, whereas reduction of ROS to background levels largely abolished TLD. We conclude that ROS contribute to TLD by converting single-stranded DNA lesions into double-stranded DNA breaks. Participation of ROS in the terminal phases of TLD provides a specific example of how ROS contribute to stress-mediated bacterial self-destruction.
Conflict of interest statement
Figures
Comment in
-
A radical way to die.Nat Microbiol. 2017 Dec;2(12):1582-1583. doi: 10.1038/s41564-017-0074-6. Nat Microbiol. 2017. PMID: 29176695 No abstract available.
References
-
- Ahmad SI, Kirk SH, Eisenstark A. Thymine metabolism and thymineless death in prokaryotes and eukaryotes. Annu. Rev. Microbiol. 1998;52:591–625. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
