

Slow molecular recognition by RNA
- PMID: 28971853
- PMCID: PMC5688996
- DOI: 10.1261/rna.062026.117
Slow molecular recognition by RNA
Abstract
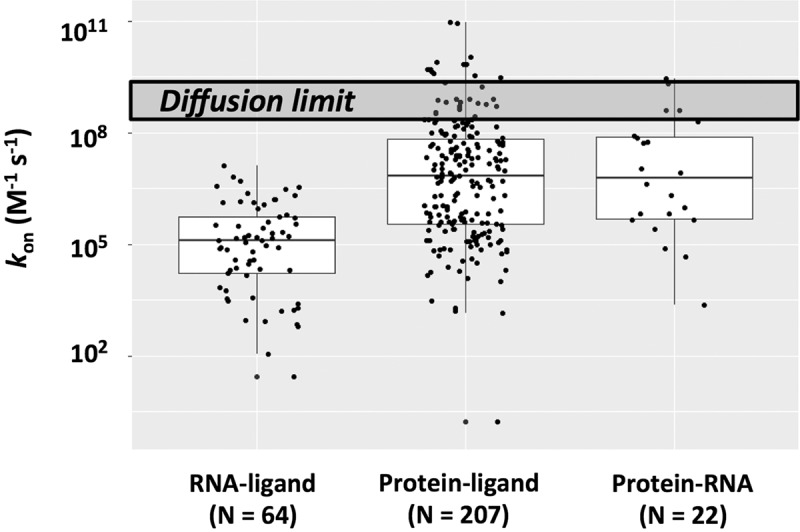
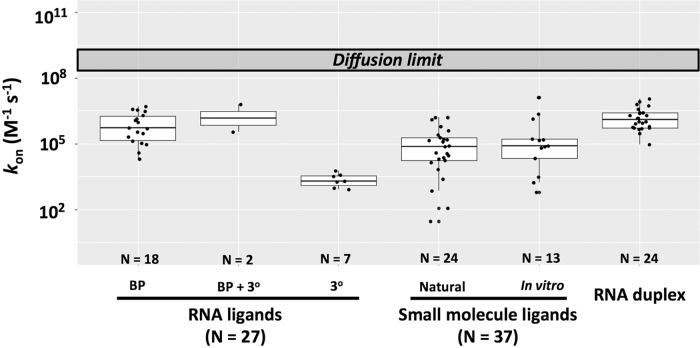
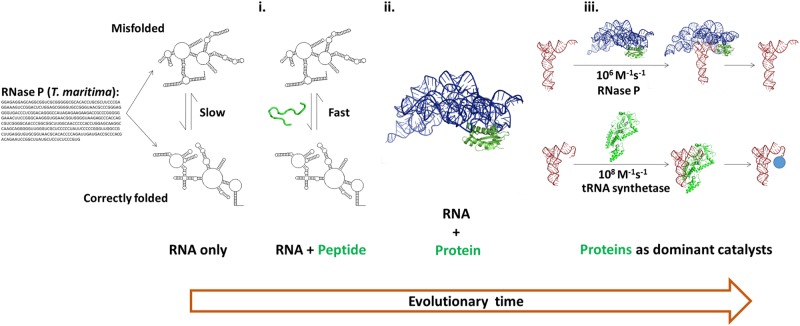
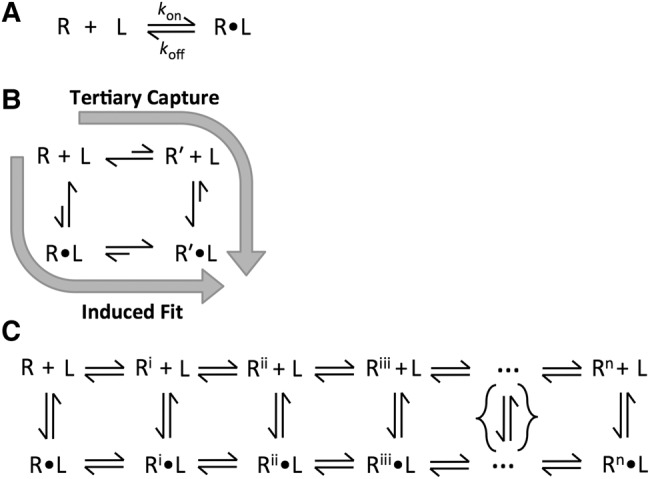
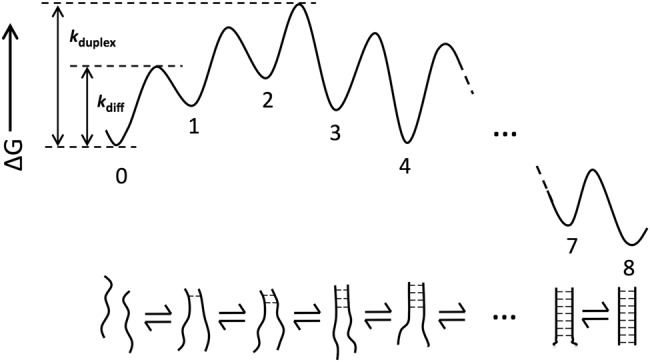
Molecular recognition is central to biological processes, function, and specificity. Proteins associate with ligands with a wide range of association rate constants, with maximal values matching the theoretical limit set by the rate of diffusional collision. As less is known about RNA association, we compiled association rate constants for all RNA/ligand complexes that we could find in the literature. Like proteins, RNAs exhibit a wide range of association rate constants. However, the fastest RNA association rates are considerably slower than those of the fastest protein associations and fall well below the diffusional limit. The apparently general observation of slow association with RNAs has implications for evolution and for modern-day biology. Our compilation highlights a quantitative molecular property that can contribute to biological understanding and underscores our need to develop a deeper physical understanding of molecular recognition events.
Keywords: RNA structure; binding kinetics; molecular recognition.
© 2017 Gleitsman et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
References
-
- Berg OG, von Hippel PH. 1985. Diffusion-controlled macromolecular interactions. Annu Rev Biophys Biophys Chem 14: 131–160. - PubMed
-
- Bloomfield VA, Crothers DM, Tinoco I. 2000. Nucleic acids: structure, properties, and functions. University Science Books, Sausalito, CA.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials