

Sustained Ex Vivo Susceptibility of Plasmodium falciparum to Artemisinin Derivatives but Increasing Tolerance to Artemisinin Combination Therapy Partner Quinolines in The Gambia
- PMID: 28971859
- PMCID: PMC5700332
- DOI: 10.1128/AAC.00759-17
Sustained Ex Vivo Susceptibility of Plasmodium falciparum to Artemisinin Derivatives but Increasing Tolerance to Artemisinin Combination Therapy Partner Quinolines in The Gambia
Abstract
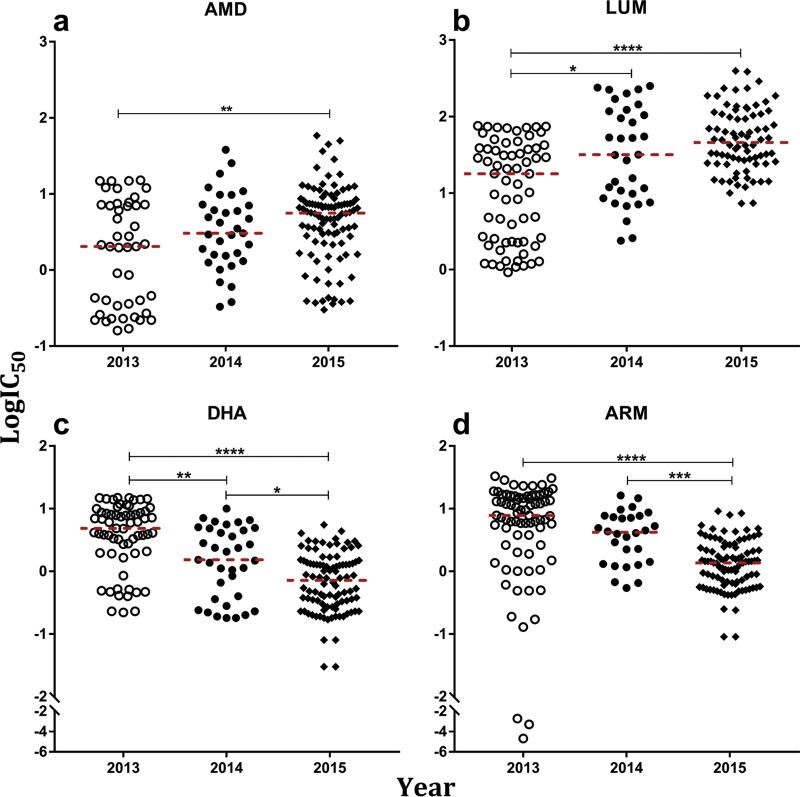
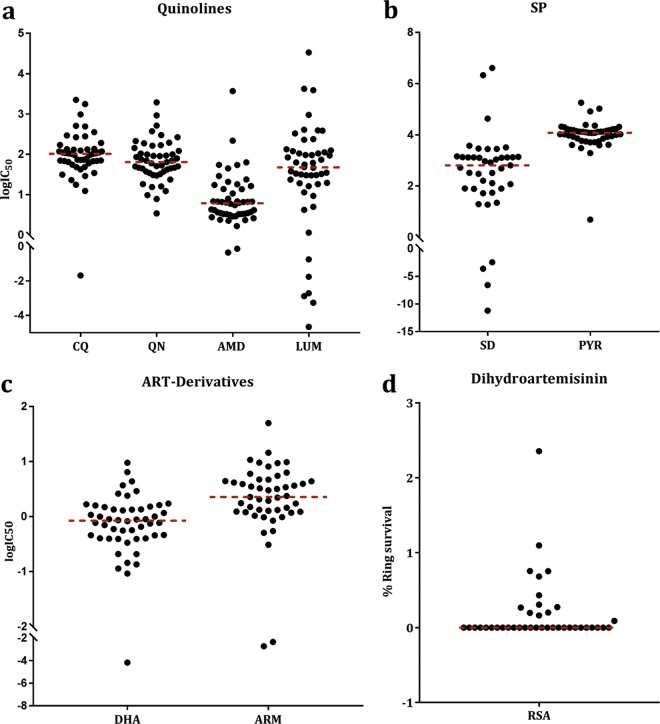
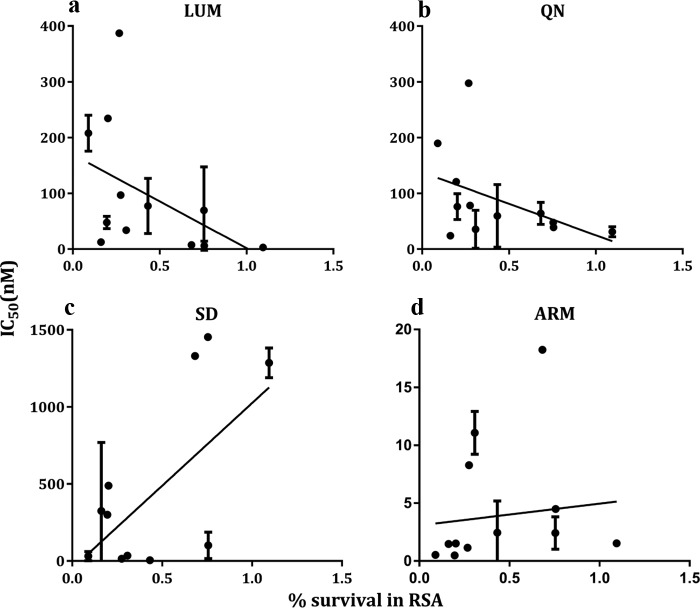
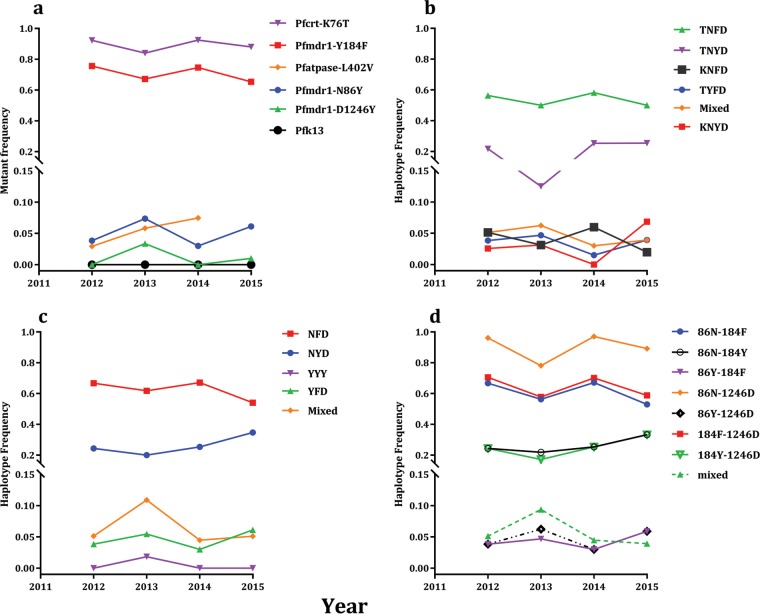
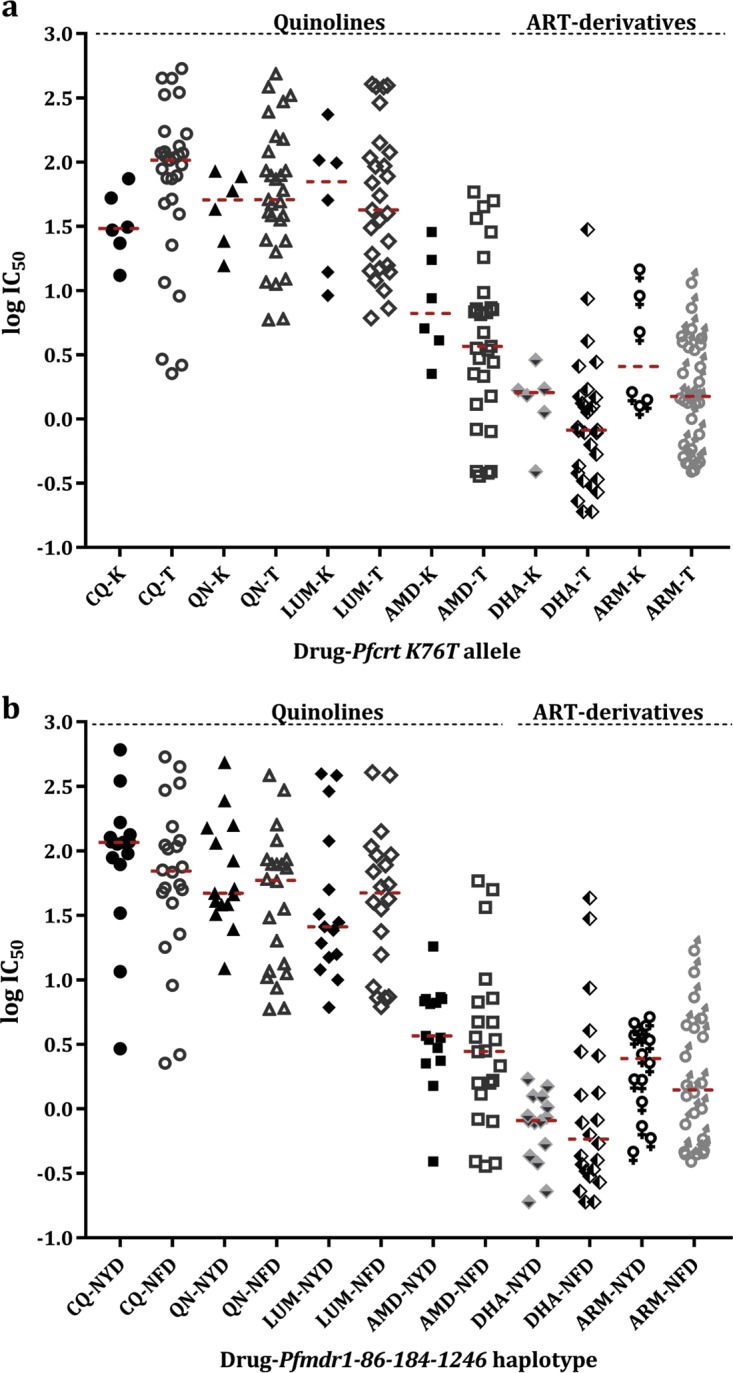
Antimalarial interventions have yielded a significant decline in malaria prevalence in The Gambia, where artemether-lumefantrine (AL) has been used as a first-line antimalarial for a decade. Clinical Plasmodium falciparum isolates collected from 2012 to 2015 were analyzed ex vivo for antimalarial susceptibility and genotyped for drug resistance markers (pfcrt K76T, pfmdr1 codons 86, 184, and 1246, and pfk13) and microsatellite variation. Additionally, allele frequencies of single nucleotide polymorphisms (SNPs) from other drug resistance-associated genes were compared from genomic sequence data sets from 2008 (n = 79) and 2014 (n = 168). No artemisinin resistance-associated pfk13 mutation was found, and only 4% of the isolates tested in 2015 showed significant growth after exposure to dihydroartemisinin. Conversely, the 50% inhibitory concentrations (IC50s) of amodiaquine and lumefantrine increased within this period. pfcrt 76T and pfmdr1 184F mutants remained at a prevalence above 80%. pfcrt 76T was positively associated with higher IC50s to chloroquine. pfmdr1 NYD increased in frequency between 2012 and 2015 due to lumefantrine selection. The TNYD (pfcrt 76T and pfmdr1 NYD wild-type haplotype) also increased in frequency following AL implementation in 2008. These results suggest selection for pfcrt and pfmdr1 genotypes that enable tolerance to lumefantrine. Increased tolerance to lumefantrine calls for sustained chemotherapeutic monitoring in The Gambia to minimize complete artemisinin combination therapy (ACT) failure in the future.
Keywords: alleles; antimalarial agents; artemisinin combination therapies; drug resistance evolution; ex vivo susceptibility; haplotypes; malaria elimination.
Copyright © 2017 Amambua-Ngwa et al.
Figures
References
-
- Greenhouse B, Slater M, Njama-Meya D, Nzarubara B, Maiteki-Sebuguzi C, Clark TD, Staedke SG, Kamya MR, Hubbard A, Rosenthal PJ, Dorsey G. 2009. Decreasing efficacy of antimalarial combination therapy in Uganda is explained by decreasing host immunity rather than increasing drug resistance. J Infect Dis 199:758–765. doi: 10.1086/596741. - DOI - PMC - PubMed
-
- Conrad MD, LeClair N, Arinaitwe E, Wanzira H, Kakuru A, Bigira V, Muhindo M, Kamya MR, Tappero JW, Greenhouse B, Dorsey G, Rosenthal PJ. 2014. Comparative impacts over 5 years of artemisinin-based combination therapies on Plasmodium falciparum polymorphisms that modulate drug sensitivity in Ugandan children. J Infect Dis 210:344–353. doi: 10.1093/infdis/jiu141. - DOI - PMC - PubMed
-
- Beshir KB, Sutherland CJ, Sawa P, Drakeley CJ, Okell L, Mweresa CK, Omar SA, Shekalaghe SA, Kaur H, Ndaro A, Chilongola J, Schallig HD, Sauerwein RW, Hallett RL, Bousema T. 2013. Residual Plasmodium falciparum parasitemia in Kenyan children after artemisinin-combination therapy is associated with increased transmission to mosquitoes and parasite recurrence. J Infect Dis 208:2017–2024. doi: 10.1093/infdis/jit431. - DOI - PMC - PubMed
-
- WWARN Artemisinin-based Combination Therapy (ACT) Africa Baseline Study Group, Dahal P, d'Alessandro U, Dorsey G, Guerin PJ, Nsanzabana C, Price RN, Sibley CH, Stepniewska K, Talisuna AO. 2015. Clinical determinants of early parasitological response to ACTs in African patients with uncomplicated falciparum malaria: a literature review and meta-analysis of individual patient data. BMC Med 13:212. doi: 10.1186/s12916-015-0445-x. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
