

Postsynaptic adhesion GPCR latrophilin-2 mediates target recognition in entorhinal-hippocampal synapse assembly
- PMID: 28972101
- PMCID: PMC5674891
- DOI: 10.1083/jcb.201703042
Postsynaptic adhesion GPCR latrophilin-2 mediates target recognition in entorhinal-hippocampal synapse assembly
Abstract
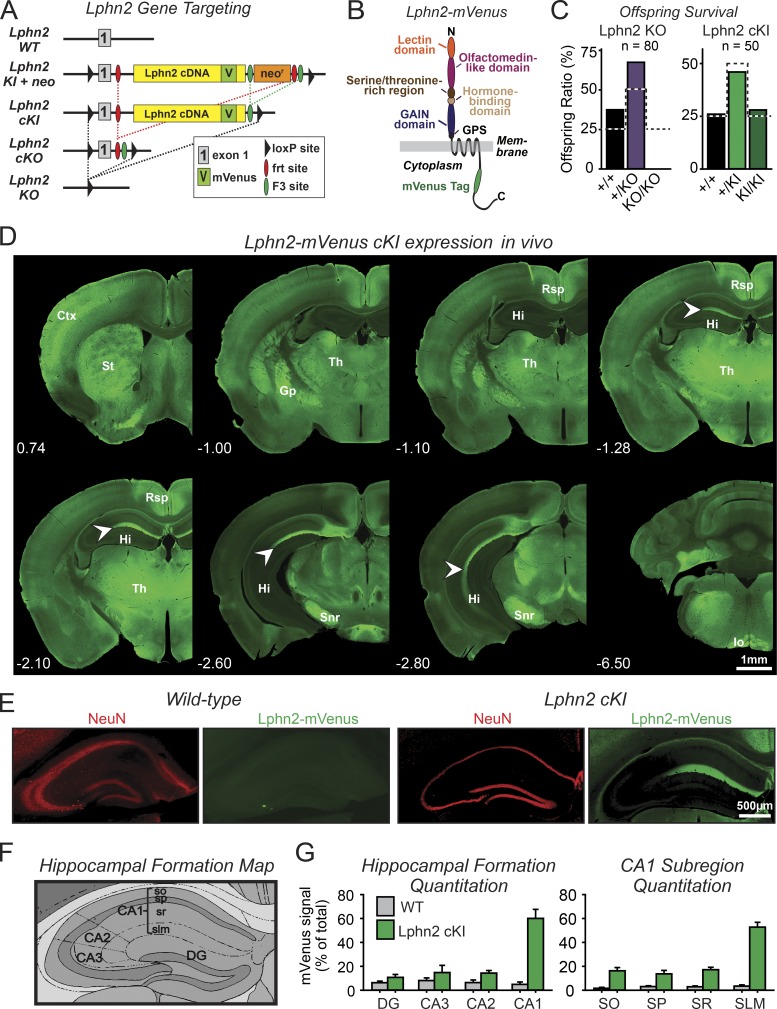
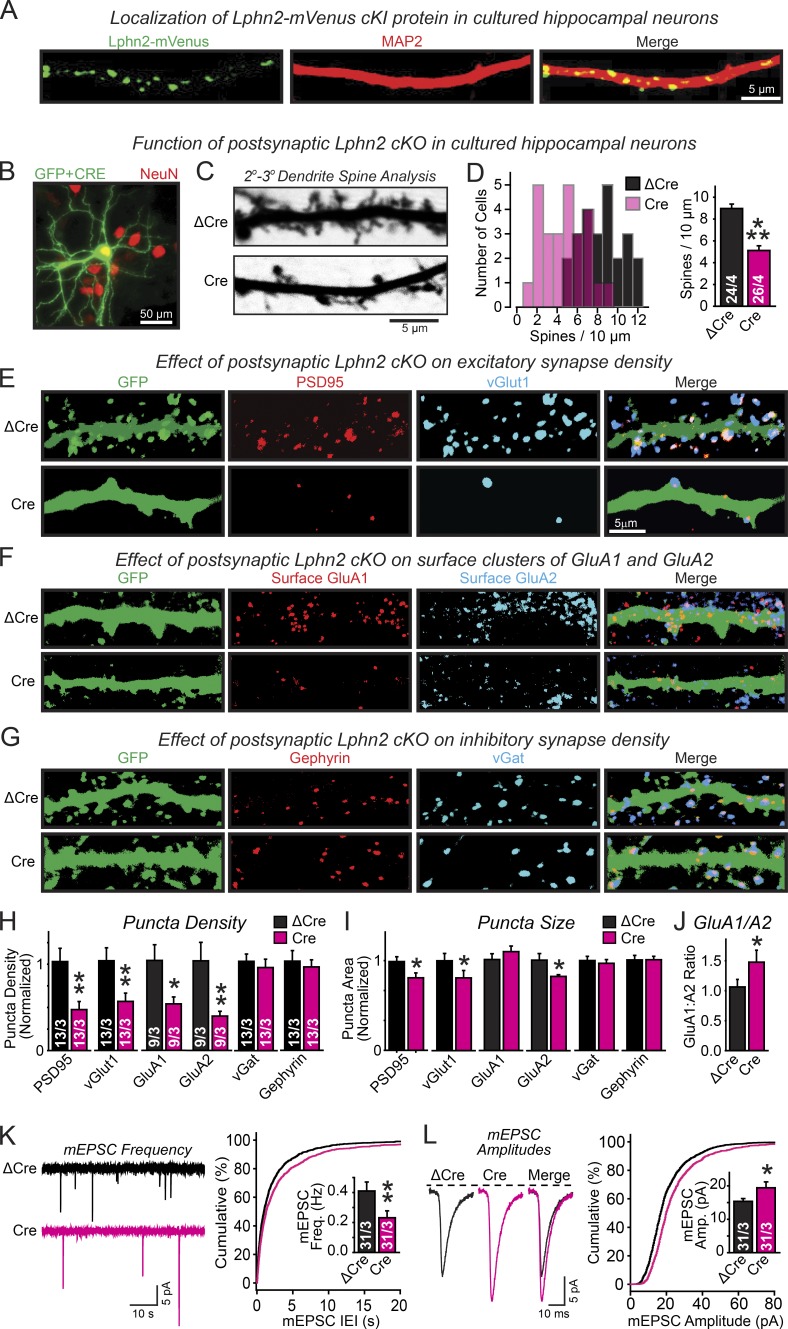
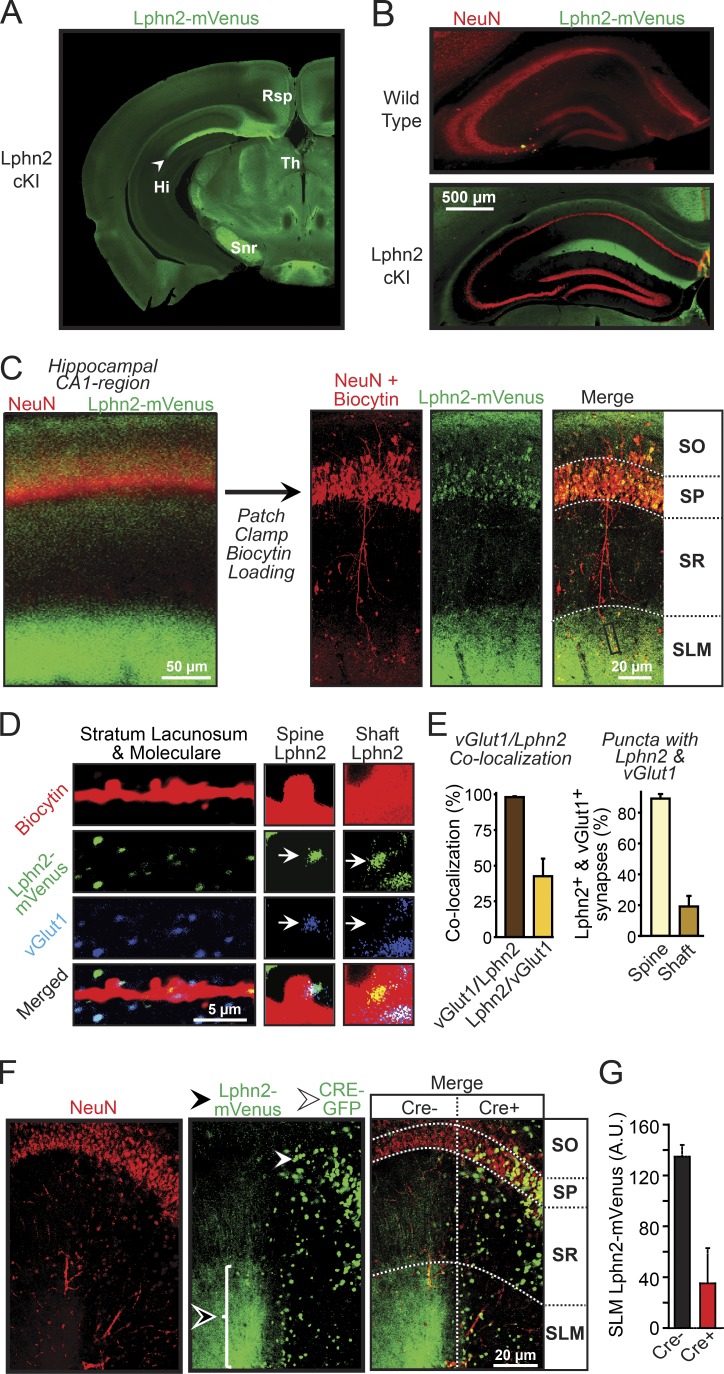
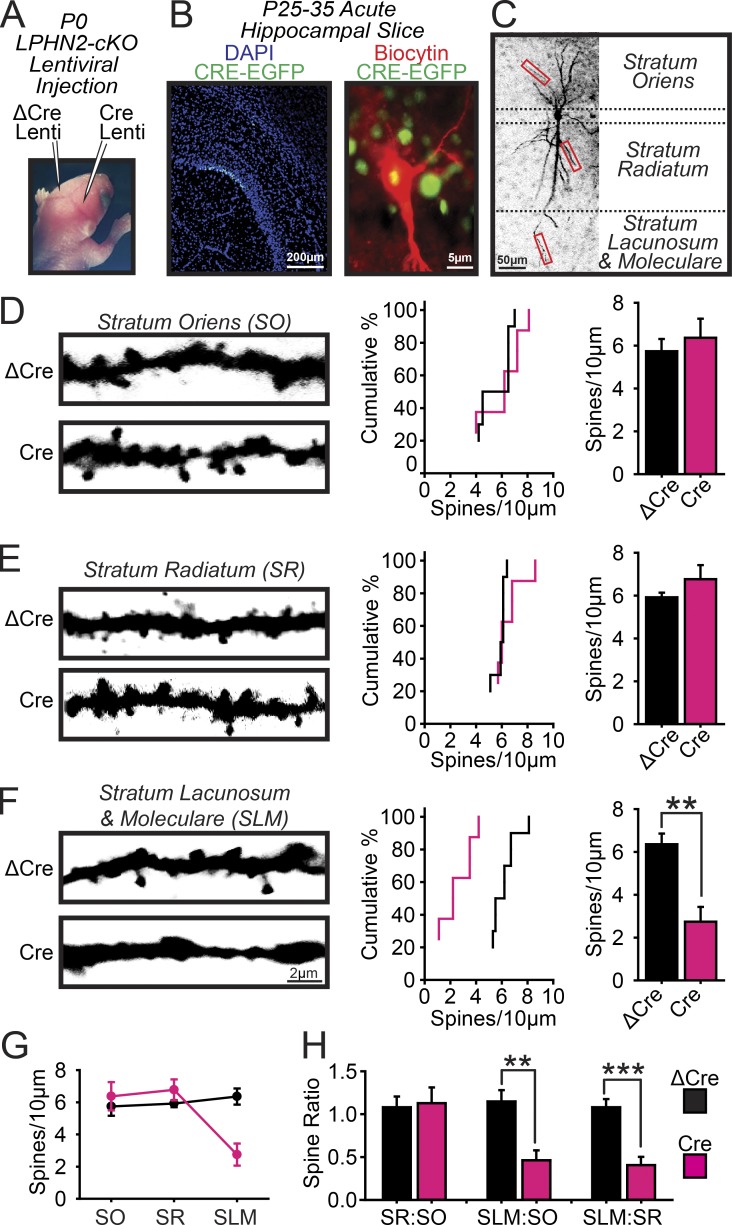
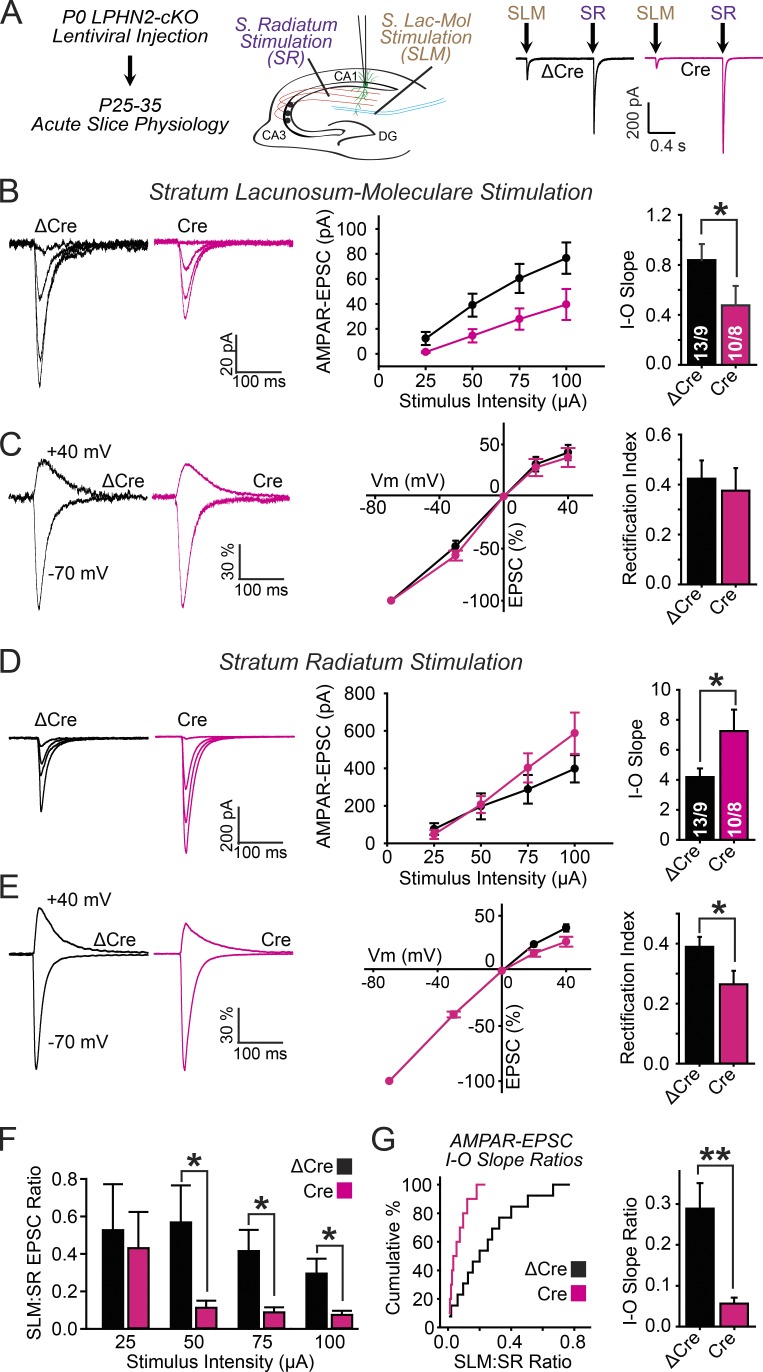
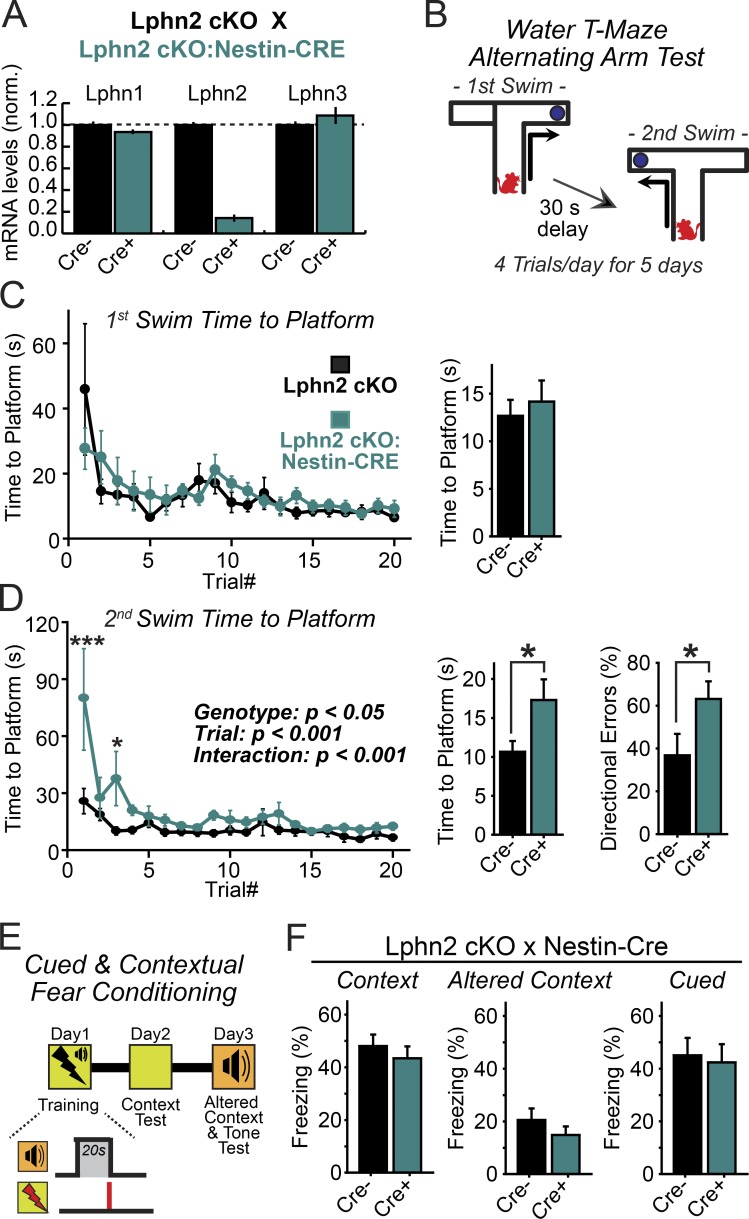
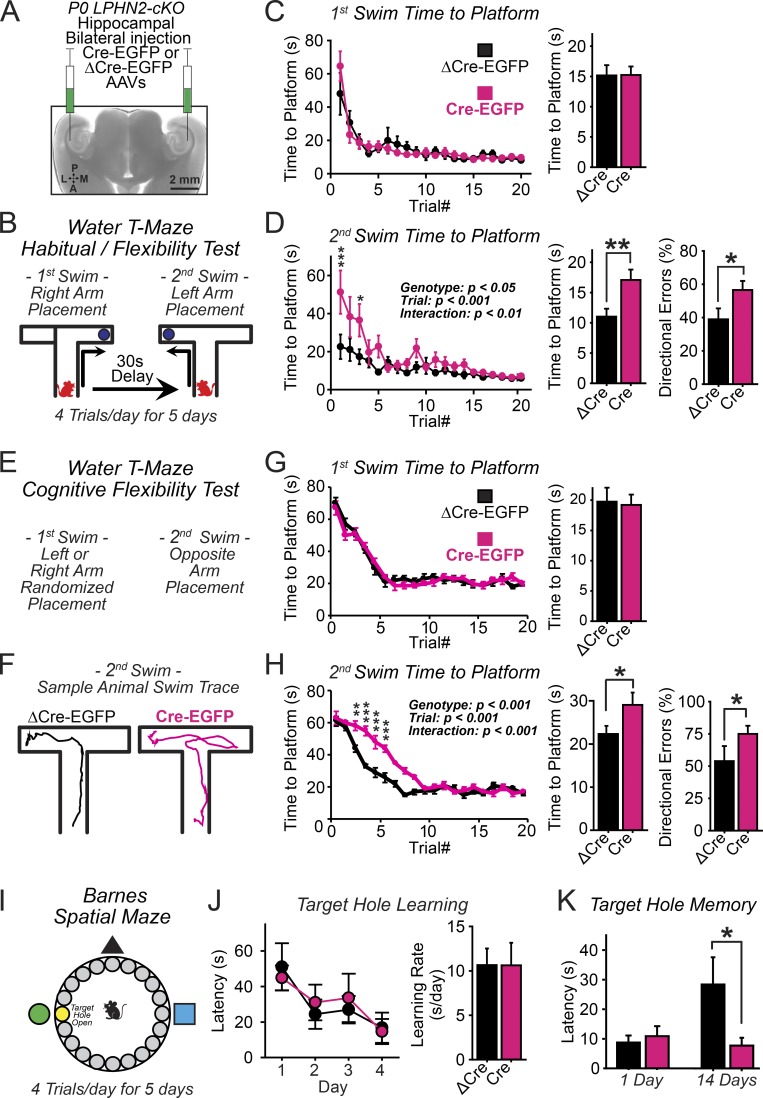
Synapse assembly likely requires postsynaptic target recognition by incoming presynaptic afferents. Using newly generated conditional knock-in and knockout mice, we show in this study that latrophilin-2 (Lphn2), a cell-adhesion G protein-coupled receptor and presumptive α-latrotoxin receptor, controls the numbers of a specific subset of synapses in CA1-region hippocampal neurons, suggesting that Lphn2 acts as a synaptic target-recognition molecule. In cultured hippocampal neurons, Lphn2 maintained synapse numbers via a postsynaptic instead of a presynaptic mechanism, which was surprising given its presumptive role as an α-latrotoxin receptor. In CA1-region neurons in vivo, Lphn2 was specifically targeted to dendritic spines in the stratum lacunosum-moleculare, which form synapses with presynaptic entorhinal cortex afferents. In this study, postsynaptic deletion of Lphn2 selectively decreased spine numbers and impaired synaptic inputs from entorhinal but not Schaffer-collateral afferents. Behaviorally, loss of Lphn2 from the CA1 region increased spatial memory retention but decreased learning of sequential spatial memory tasks. Thus, Lphn2 appears to control synapse numbers in the entorhinal cortex/CA1 region circuit by acting as a domain-specific postsynaptic target-recognition molecule.
© 2017 Anderson et al.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
