

Fremanezumab-A Humanized Monoclonal Anti-CGRP Antibody-Inhibits Thinly Myelinated (Aδ) But Not Unmyelinated (C) Meningeal Nociceptors
- PMID: 28972120
- PMCID: PMC5666582
- DOI: 10.1523/JNEUROSCI.2211-17.2017
Fremanezumab-A Humanized Monoclonal Anti-CGRP Antibody-Inhibits Thinly Myelinated (Aδ) But Not Unmyelinated (C) Meningeal Nociceptors
Abstract
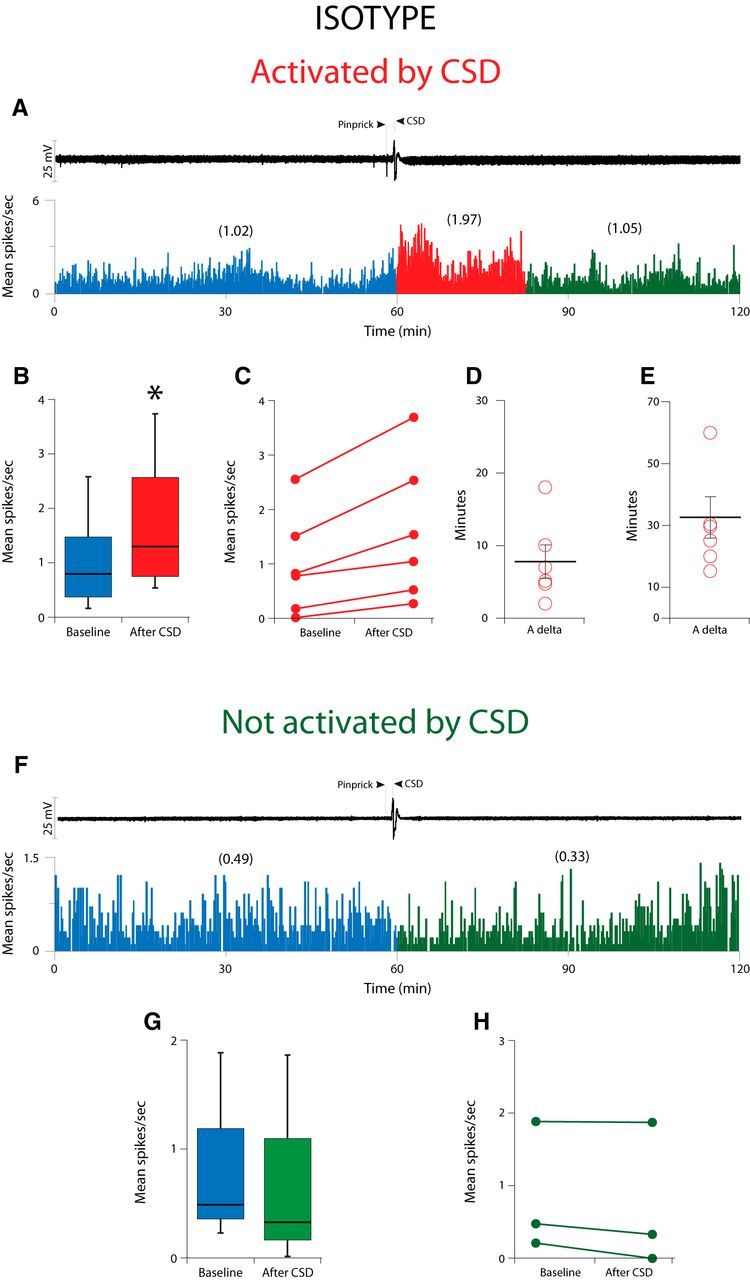
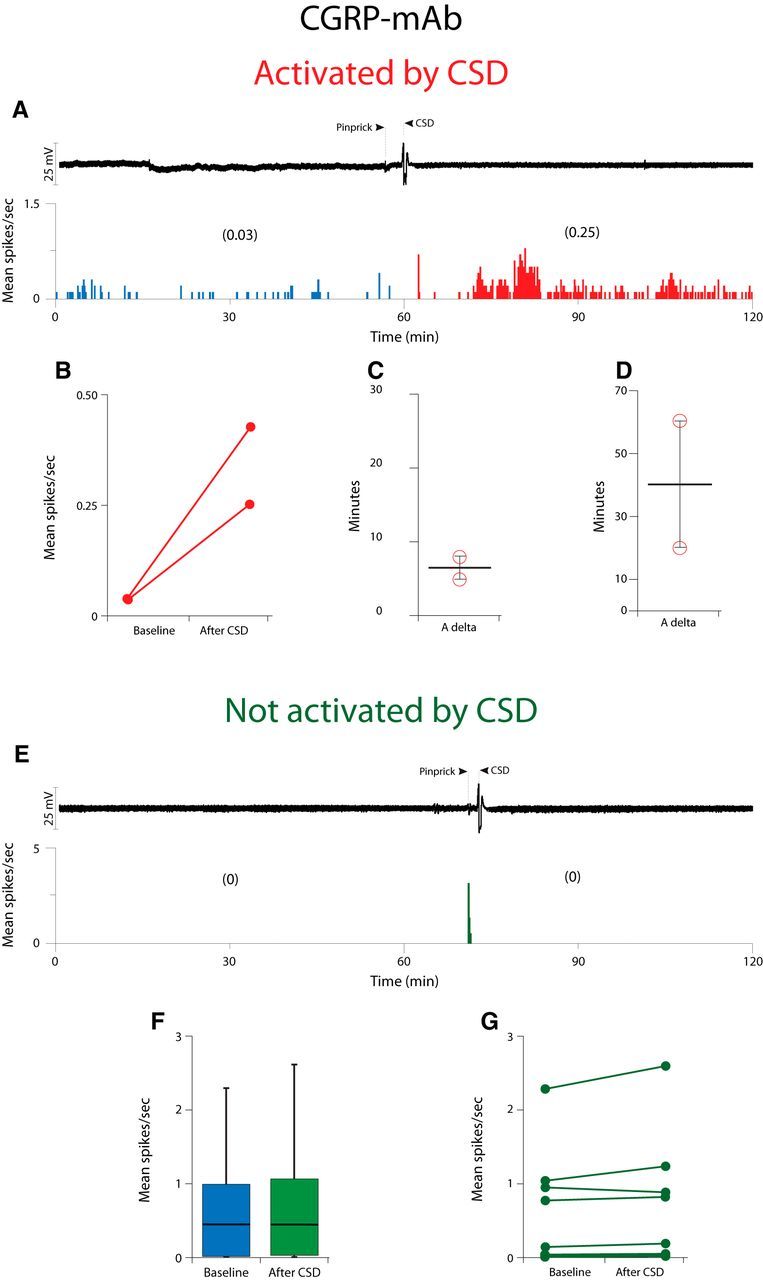
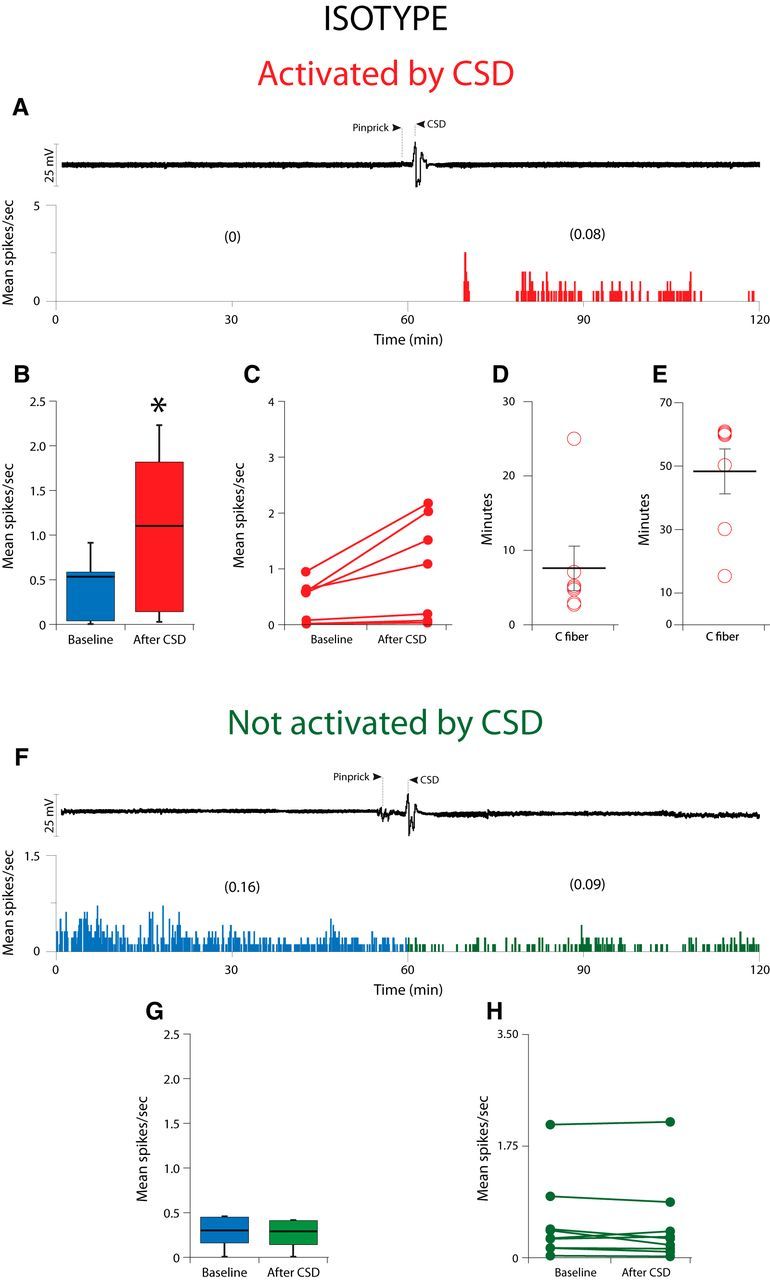
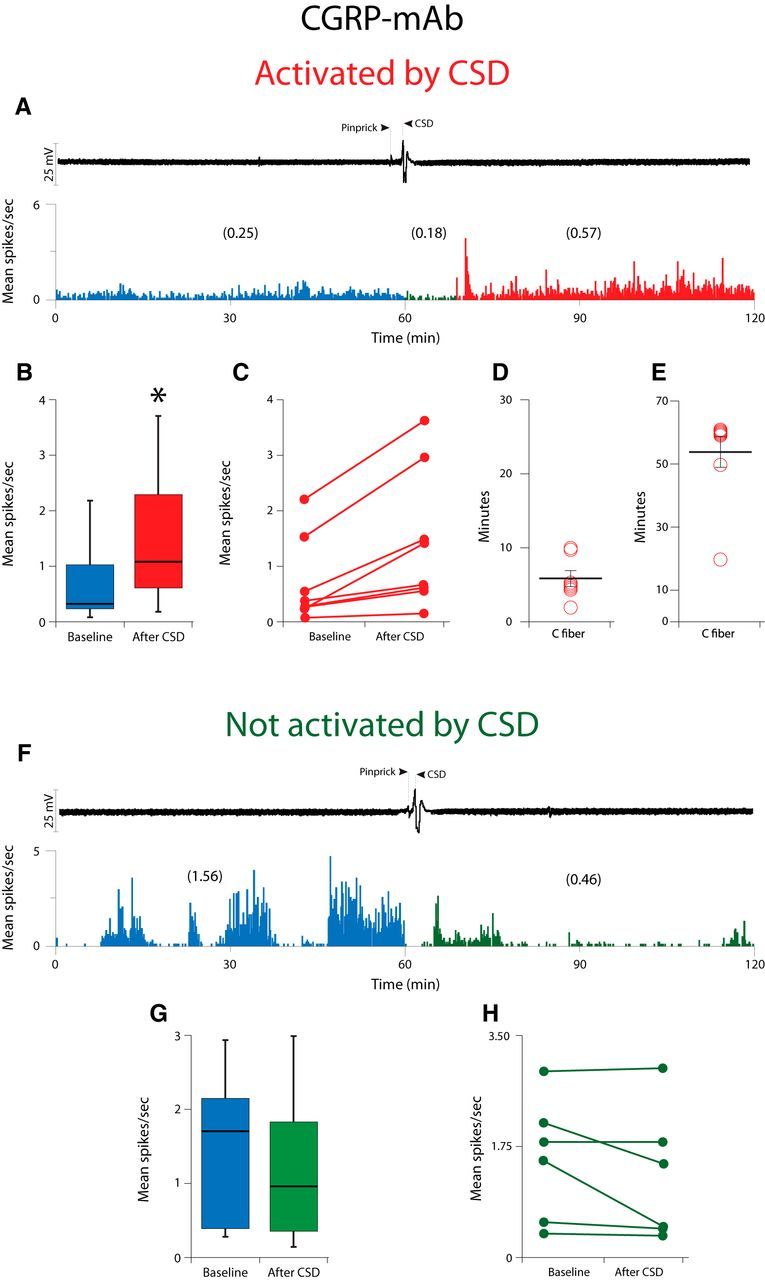
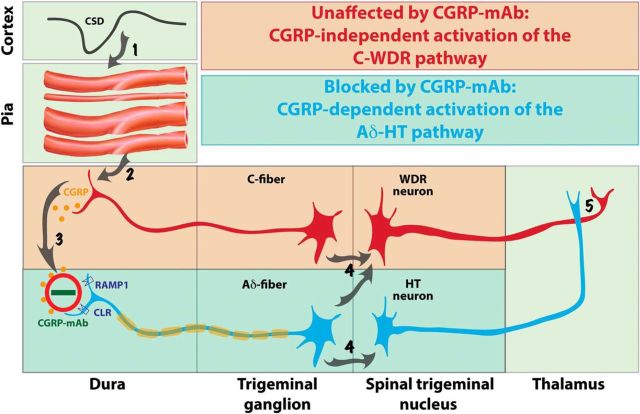
Calcitonin gene-related peptide (CGRP), the most abundant neuropeptide in primary afferent sensory neurons, is strongly implicated in the pathophysiology of migraine headache, but its role in migraine is still equivocal. As a new approach to migraine treatment, humanized anti-CGRP monoclonal antibodies (CGRP-mAbs) were developed to reduce the availability of CGRP, and were found effective in reducing the frequency of chronic and episodic migraine. We recently tested the effect of fremanezumab (TEV-48125), a CGRP-mAb, on the activity of second-order trigeminovascular dorsal horn neurons that receive peripheral input from the cranial dura, and found a selective inhibition of high-threshold but not wide-dynamic range class of neurons. To investigate the basis for this selective inhibitory effect, and further explore the mechanism of action of CGRP-mAbs, we tested the effect of fremanezumab on the cortical spreading depression-evoked activation of mechanosensitive primary afferent meningeal nociceptors that innervate the cranial dura, using single-unit recording in the trigeminal ganglion of anesthetized male rats. Fremanezumab pretreatment selectively inhibited the responsiveness of Aδ neurons, but not C-fiber neurons, as reflected in a decrease in the percentage of neurons that showed activation by cortical spreading depression. These findings identify Aδ meningeal nociceptors as a likely site of action of fremanezumab in the prevention of headache. The selectivity in its peripheral inhibitory action may partly account for fremanezumab's selective inhibition of high-threshold, as a result of a predominant A-δ input to high-threshold neurons, but not wide dynamic-range dorsal horn neurons, and why it may not be effective in all migraine patients.SIGNIFICANCE STATEMENT Recently, we reported that humanized CGRP monoclonal antibodies (CGRP-mAbs) prevent activation and sensitization of high-threshold (HT) but not wide-dynamic range trigeminovascular neurons by cortical spreading depression (CSD). In the current paper, we report that CGRP-mAbs prevent the activation of Aδ but not C-type meningeal nociceptors by CSD. This is the first identification of an anti-migraine drug that appears to be selective for Aδ-fibers (peripherally) and HT neurons (centrally). As the main CGRP-mAb site of action appears to be situated outside the brain, we conclude that the initiation of the headache phase of migraine depends on activation of meningeal nociceptors, and that for selected patients, activation of the Aδ-HT pain pathway may be sufficient for the generation of headache perception.
Keywords: calcitonin gene-related peptide; cortical spreading depression; headache; migraine; pain; trigeminovascular.
Copyright © 2017 the authors 0270-6474/17/3710587-10$15.00/0.
Figures
Similar articles
-
Selective Inhibition of Trigeminovascular Neurons by Fremanezumab: A Humanized Monoclonal Anti-CGRP Antibody.J Neurosci. 2017 Jul 26;37(30):7149-7163. doi: 10.1523/JNEUROSCI.0576-17.2017. Epub 2017 Jun 22. J Neurosci. 2017. PMID: 28642283 Free PMC article.
-
Atogepant - an orally-administered CGRP antagonist - attenuates activation of meningeal nociceptors by CSD.Cephalalgia. 2022 Aug;42(9):933-943. doi: 10.1177/03331024221083544. Epub 2022 Mar 25. Cephalalgia. 2022. PMID: 35332801 Free PMC article.
-
Novel insight into atogepant mechanisms of action in migraine prevention.Brain. 2024 Aug 1;147(8):2884-2896. doi: 10.1093/brain/awae062. Brain. 2024. PMID: 38411458 Free PMC article.
-
Pathophysiological Mechanisms in Migraine and the Identification of New Therapeutic Targets.CNS Drugs. 2019 Jun;33(6):525-537. doi: 10.1007/s40263-019-00630-6. CNS Drugs. 2019. PMID: 30989485 Review.
-
Fremanezumab for the preventive treatment of migraine in adults.Expert Rev Clin Pharmacol. 2019 Aug;12(8):741-748. doi: 10.1080/17512433.2019.1635452. Epub 2019 Jun 26. Expert Rev Clin Pharmacol. 2019. PMID: 31220963 Review.
Cited by
-
Onabotulinumtoxin-A: Previous Prophylactic Treatment Might Improve Subsequent Anti-CGRP Monoclonal Antibodies Response in Patients with Chronic Migraine.Toxins (Basel). 2023 Nov 30;15(12):677. doi: 10.3390/toxins15120677. Toxins (Basel). 2023. PMID: 38133181 Free PMC article.
-
Current understanding of osteoarthritis pathogenesis and relevant new approaches.Bone Res. 2022 Sep 20;10(1):60. doi: 10.1038/s41413-022-00226-9. Bone Res. 2022. PMID: 36127328 Free PMC article. Review.
-
Fluorescently-labeled fremanezumab is distributed to sensory and autonomic ganglia and the dura but not to the brain of rats with uncompromised blood brain barrier.Cephalalgia. 2020 Mar;40(3):229-240. doi: 10.1177/0333102419896760. Epub 2019 Dec 19. Cephalalgia. 2020. PMID: 31856583 Free PMC article.
-
Effect of Xiongmatang Extract on Behavioral and TRPV1-CGRP/CGRP-R Pathway in Rats With Migraine.Front Pharmacol. 2022 Mar 8;13:835187. doi: 10.3389/fphar.2022.835187. eCollection 2022. Front Pharmacol. 2022. PMID: 35350752 Free PMC article. Review.
-
CGRP in Animal Models of Migraine.Handb Exp Pharmacol. 2019;255:85-107. doi: 10.1007/164_2018_187. Handb Exp Pharmacol. 2019. PMID: 30689086 Free PMC article. Review.
References
-
- Bigal ME, Dodick DW, Rapoport AM, Silberstein SD, Ma Y, Yang R, Loupe PS, Burstein R, Newman LC, Lipton RB (2015a) Safety, tolerability, and efficacy of TEV-48125 for preventive treatment of high-frequency episodic migraine: a multicentre, randomised, double-blind, placebo-controlled, phase 2b study. Lancet Neurol 14:1081–1090. 10.1016/S1474-4422(15)00249-5 - DOI - PubMed
-
- Bigal ME, Edvinsson L, Rapoport AM, Lipton RB, Spierings EL, Diener HC, Burstein R, Loupe PS, Ma Y, Yang R, Silberstein SD (2015b) Safety, tolerability, and efficacy of TEV-48125 for preventive treatment of chronic migraine: a multicentre, randomised, double-blind, placebo-controlled, phase 2b study. Lancet Neurol 14:1091–1100. 10.1016/S1474-4422(15)00245-8 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials