

PelA and PelB proteins form a modification and secretion complex essential for Pel polysaccharide-dependent biofilm formation in Pseudomonas aeruginosa
- PMID: 28972168
- PMCID: PMC5702679
- DOI: 10.1074/jbc.M117.812842
PelA and PelB proteins form a modification and secretion complex essential for Pel polysaccharide-dependent biofilm formation in Pseudomonas aeruginosa
Abstract
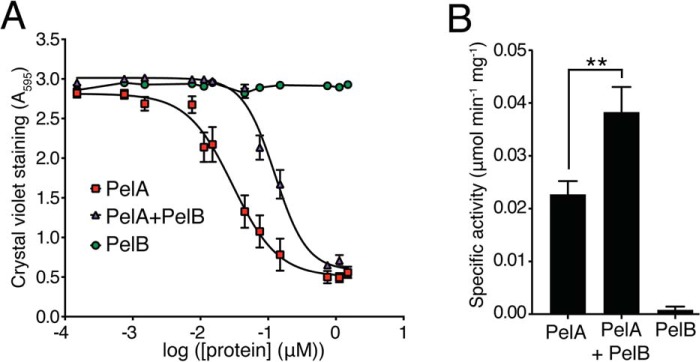
The pellicle (PEL) polysaccharide is synthesized by the opportunistic pathogen Pseudomonas aeruginosa and is an important biofilm constituent critical for bacterial virulence and persistence. PEL is a cationic polymer that promotes cell-cell interactions within the biofilm matrix through electrostatic interactions with extracellular DNA. Translocation of PEL across the outer membrane is proposed to occur via PelB, a membrane-embedded porin with a large periplasmic domain predicted to contain 19 tetratricopeptide repeats (TPRs). TPR-containing domains are typically involved in protein-protein interactions, and we therefore sought to determine whether PelB serves as a periplasmic scaffold that recruits other components of the PEL secretion apparatus. In this study, we show that the TPR domain of PelB interacts with PelA, an enzyme with PEL deacetylase and hydrolase activities. Structure determination of PelB TPRs 8-11 enabled us to design systematic deletions of individual TPRs and revealed that repeats 9-14, which are required for the cellular localization of PelA with PelB are also essential for PEL-dependent biofilm formation. Copurification experiments indicated that the interaction between PelA and PelB is direct and that the deacetylase activity of PelA increases and its hydrolase activity decreases when these proteins interact. Combined, our results indicate that the TPR-containing domain of PelB localizes PelA to the PEL secretion apparatus within the periplasm and that this may allow for efficient deacetylation of PEL before its export from the cell.
Keywords: Pseudomonas aeruginosa (P. aeruginosa); X-ray crystallography; biofilm; polysaccharide; protein-protein interaction.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures





Similar articles
-
Structural and functional analysis of Pseudomonas aeruginosa PelA provides insight into the modification of the Pel exopolysaccharide.J Biol Chem. 2025 May;301(5):108432. doi: 10.1016/j.jbc.2025.108432. Epub 2025 Mar 20. J Biol Chem. 2025. PMID: 40120681 Free PMC article.
-
Marine-Derived Cytosine Arabinoside (Ara-C) Inhibits Biofilm Formation by Inhibiting PEL Operon Proteins (Pel A and Pel B) of Pseudomonas aeruginosa: An In Silico Approach.Mol Biotechnol. 2025 May;67(5):1924-1938. doi: 10.1007/s12033-024-01169-8. Epub 2024 May 13. Mol Biotechnol. 2025. PMID: 38739212
-
PelA deacetylase activity is required for Pel polysaccharide synthesis in Pseudomonas aeruginosa.J Bacteriol. 2013 May;195(10):2329-39. doi: 10.1128/JB.02150-12. Epub 2013 Mar 15. J Bacteriol. 2013. PMID: 23504011 Free PMC article.
-
Role of polysaccharides in Pseudomonas aeruginosa biofilm development.Curr Opin Microbiol. 2007 Dec;10(6):644-8. doi: 10.1016/j.mib.2007.09.010. Epub 2007 Nov 5. Curr Opin Microbiol. 2007. PMID: 17981495 Free PMC article. Review.
-
Pseudomonas aeruginosa biofilm exopolysaccharides: assembly, function, and degradation.FEMS Microbiol Rev. 2023 Nov 1;47(6):fuad060. doi: 10.1093/femsre/fuad060. FEMS Microbiol Rev. 2023. PMID: 37884397 Free PMC article. Review.
Cited by
-
Identification of Compounds Preventing A. fumigatus Biofilm Formation by Inhibition of the Galactosaminogalactan Deacetylase Agd3.Int J Mol Sci. 2023 Jan 17;24(3):1851. doi: 10.3390/ijms24031851. Int J Mol Sci. 2023. PMID: 36768176 Free PMC article.
-
Pel Polysaccharide Biosynthesis Requires an Inner Membrane Complex Comprised of PelD, PelE, PelF, and PelG.J Bacteriol. 2020 Mar 26;202(8):e00684-19. doi: 10.1128/JB.00684-19. Print 2020 Mar 26. J Bacteriol. 2020. PMID: 31988082 Free PMC article.
-
Assembly and the gating mechanism of the Pel exopolysaccharide export complex PelBC of Pseudomonas aeruginosa.Nat Commun. 2025 Jun 5;16(1):5249. doi: 10.1038/s41467-025-60605-8. Nat Commun. 2025. PMID: 40473691 Free PMC article.
-
Untethering and Degradation of the Polysaccharide Matrix Are Essential Steps in the Dispersion Response of Pseudomonas aeruginosa Biofilms.J Bacteriol. 2020 Jan 15;202(3):e00575-19. doi: 10.1128/JB.00575-19. Print 2020 Jan 15. J Bacteriol. 2020. PMID: 31712279 Free PMC article.
-
Treatment with paeoniflorin increases lifespan of Pseudomonas aeruginosa infected Caenorhabditis elegans by inhibiting bacterial accumulation in intestinal lumen and biofilm formation.Front Pharmacol. 2023 Mar 27;14:1114219. doi: 10.3389/fphar.2023.1114219. eCollection 2023. Front Pharmacol. 2023. PMID: 37050896 Free PMC article.
References
-
- Friedman L., and Kolter R. (2004) Genes involved in matrix formation in Pseudomonas aeruginosa PA14 biofilms. Mol. Microbiol. 51, 675–690 - PubMed
-
- Ramsey D. M., and Wozniak D. J. (2005) Understanding the control of Pseudomonas aeruginosa alginate synthesis and the prospects for management of chronic infections in cystic fibrosis. Mol. Microbiol. 56, 309–322 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
