

Calmodulin regulates Cav3 T-type channels at their gating brake
- PMID: 28972185
- PMCID: PMC5723990
- DOI: 10.1074/jbc.M117.807925
Calmodulin regulates Cav3 T-type channels at their gating brake
Abstract
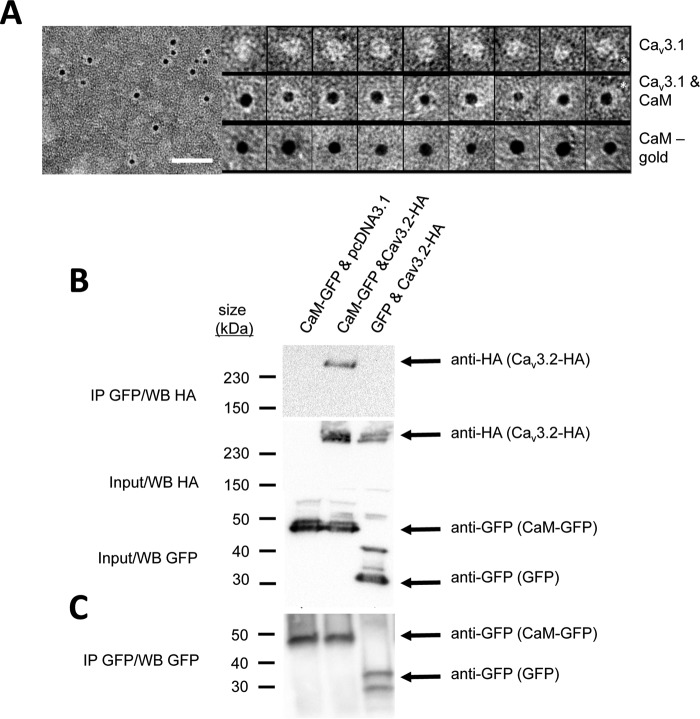
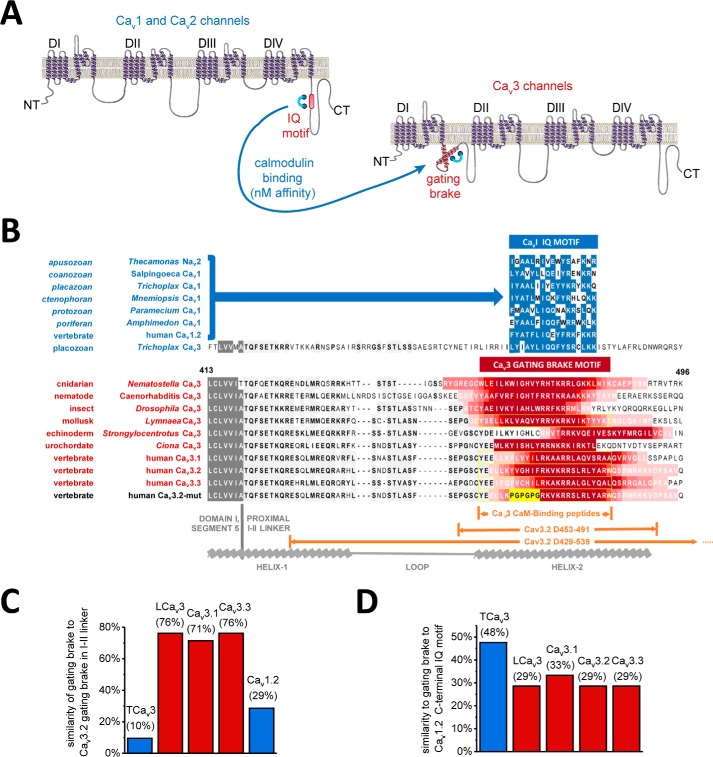
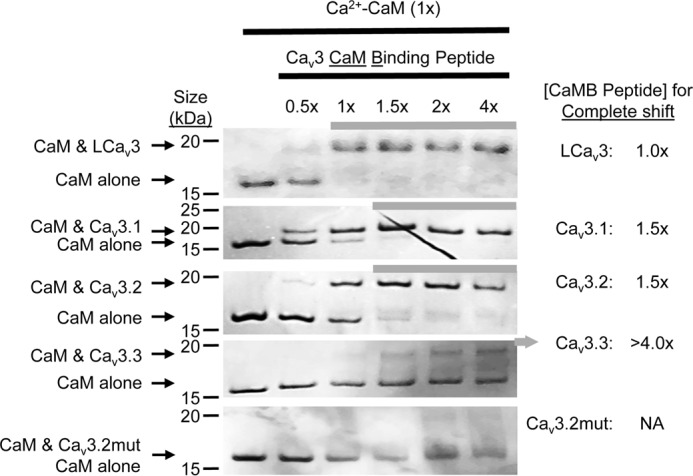
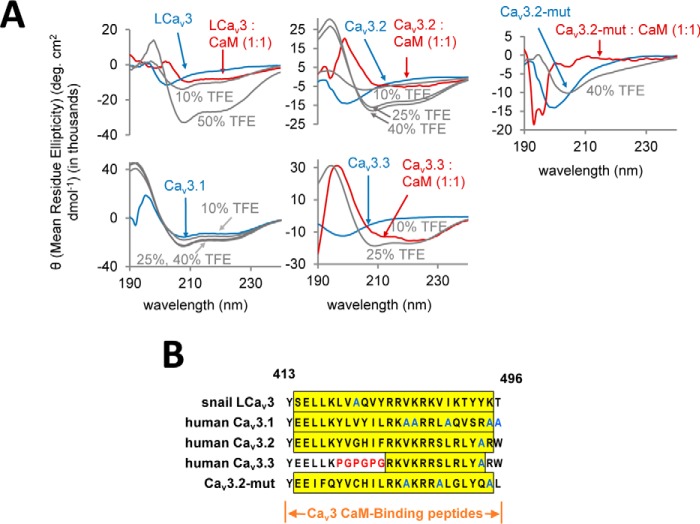
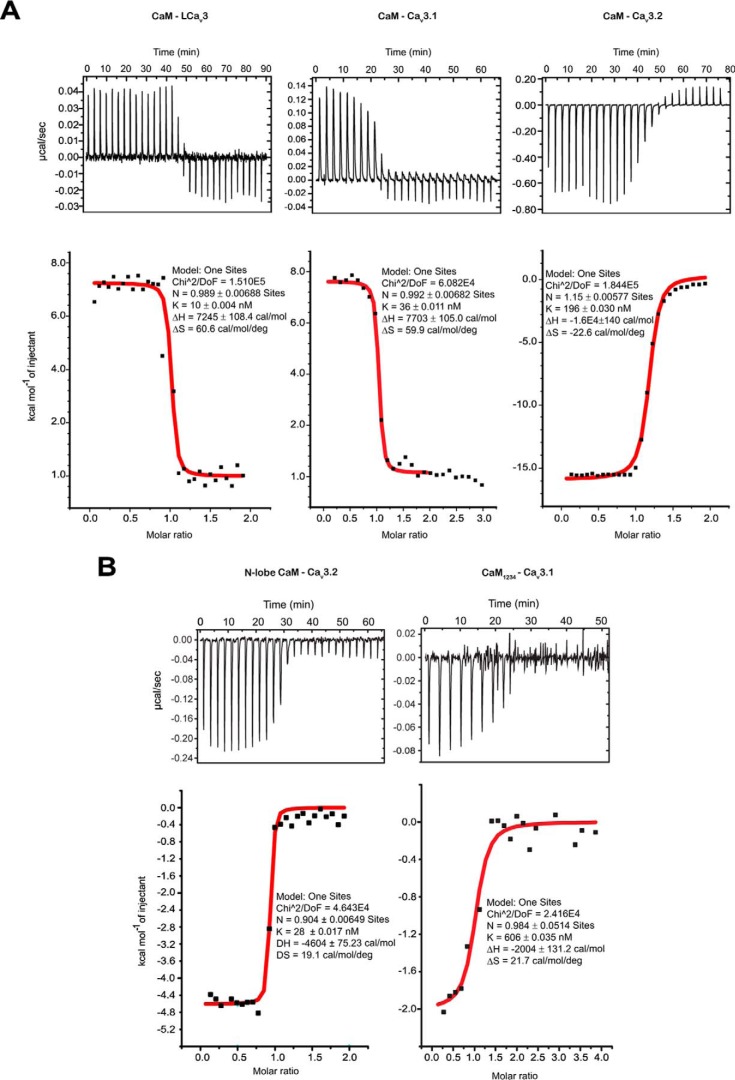
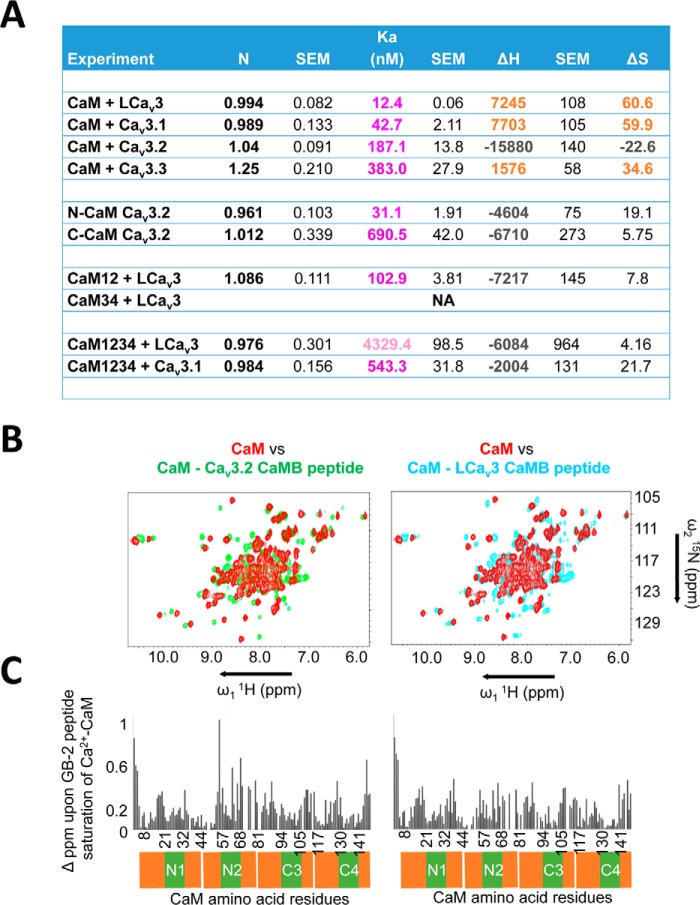
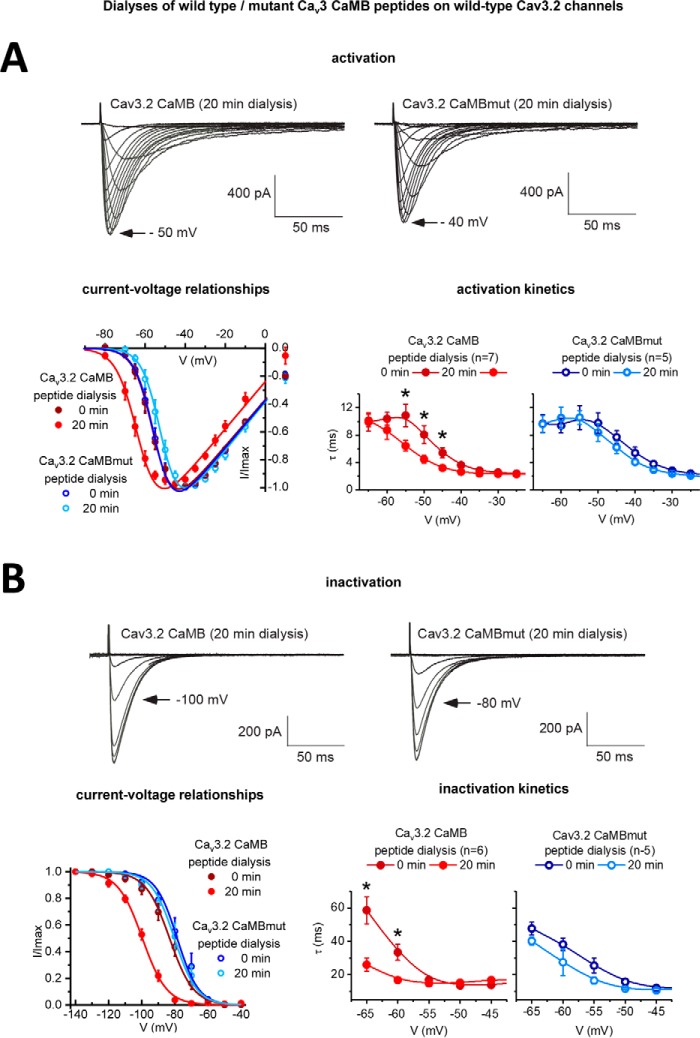
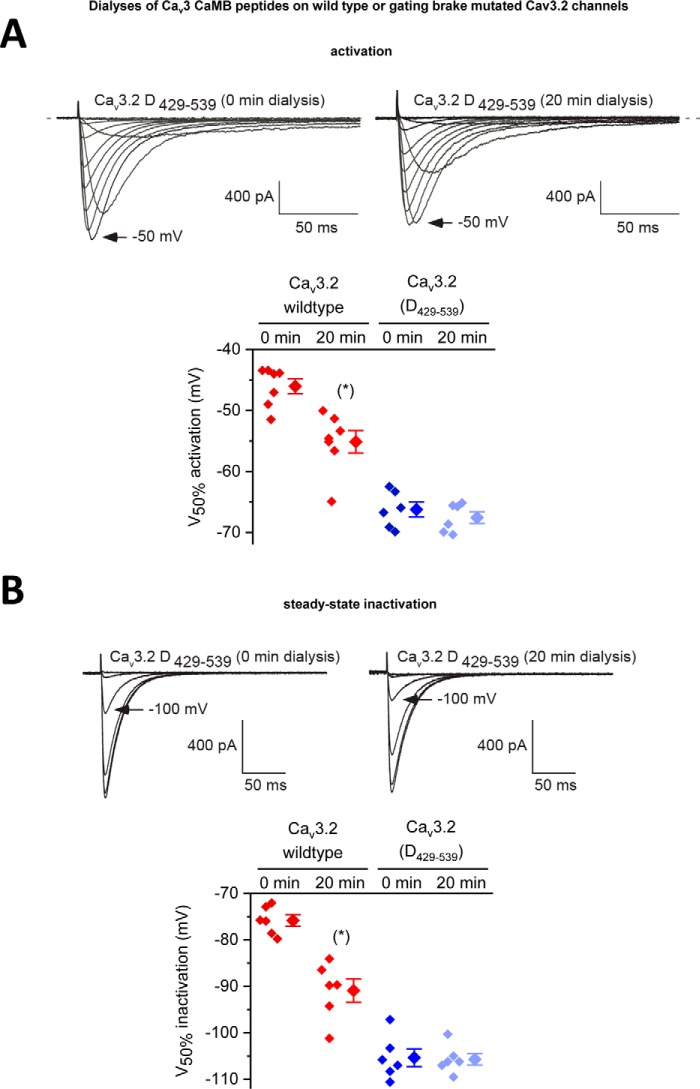
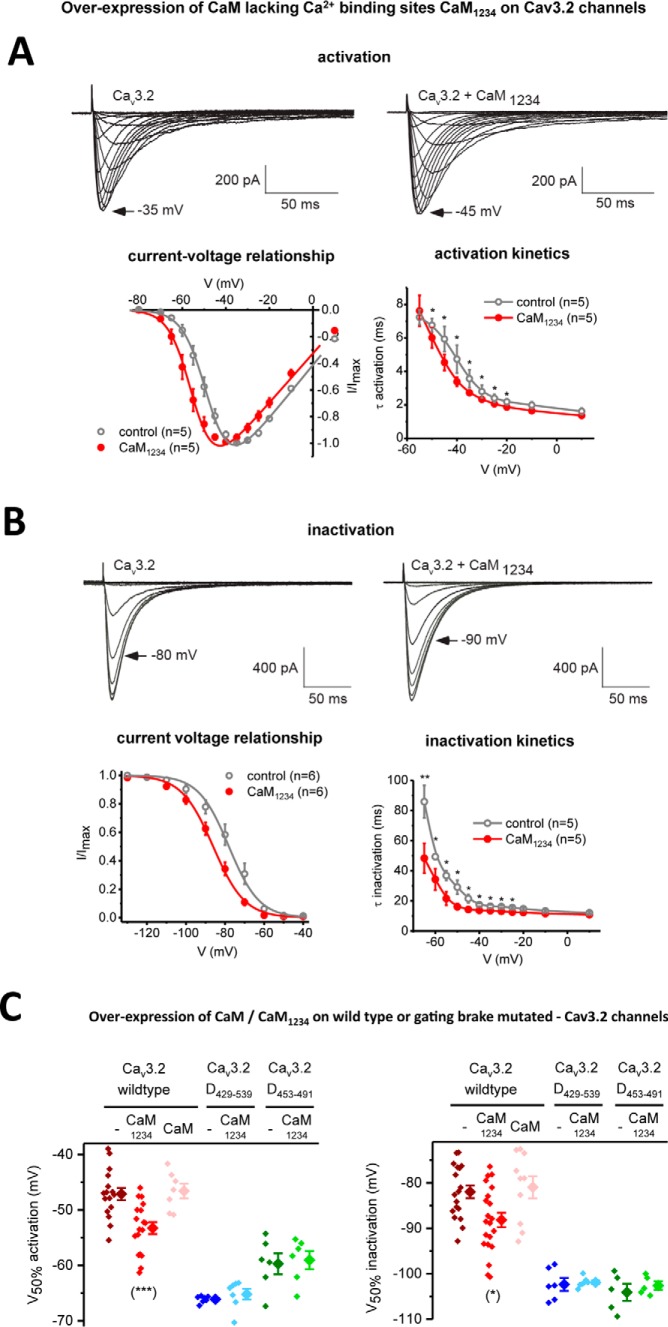
Calcium (Cav1 and Cav2) and sodium channels possess homologous CaM-binding motifs, known as IQ motifs in their C termini, which associate with calmodulin (CaM), a universal calcium sensor. Cav3 T-type channels, which serve as pacemakers of the mammalian brain and heart, lack a C-terminal IQ motif. We illustrate that T-type channels associate with CaM using co-immunoprecipitation experiments and single particle cryo-electron microscopy. We demonstrate that protostome invertebrate (LCav3) and human Cav3.1, Cav3.2, and Cav3.3 T-type channels specifically associate with CaM at helix 2 of the gating brake in the I-II linker of the channels. Isothermal titration calorimetry results revealed that the gating brake and CaM bind each other with high-nanomolar affinity. We show that the gating brake assumes a helical conformation upon binding CaM, with associated conformational changes to both CaM lobes as indicated by amide chemical shifts of the amino acids of CaM in 1H-15N HSQC NMR spectra. Intact Ca2+-binding sites on CaM and an intact gating brake sequence (first 39 amino acids of the I-II linker) were required in Cav3.2 channels to prevent the runaway gating phenotype, a hyperpolarizing shift in voltage sensitivities and faster gating kinetics. We conclude that the presence of high-nanomolar affinity binding sites for CaM at its universal gating brake and its unique form of regulation via the tuning of the voltage range of activity could influence the participation of Cav3 T-type channels in heart and brain rhythms. Our findings may have implications for arrhythmia disorders arising from mutations in the gating brake or CaM.
Keywords: calcium channel; circular dichroism (CD); cryo-electron microscopy; gating; isothermal titration calorimetry (ITC); nuclear magnetic resonance (NMR); patch clamp; short hairpin RNA (shRNA).
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
Characterization of the gating brake in the I-II loop of CaV3 T-type calcium channels.Channels (Austin). 2010 Nov-Dec;4(6):453-8. doi: 10.4161/chan.4.6.12889. Epub 2010 Nov 1. Channels (Austin). 2010. PMID: 21099341 Free PMC article. Review.
-
Contrasting the roles of the I-II loop gating brake in CaV3.1 and CaV3.3 calcium channels.Pflugers Arch. 2015 Dec;467(12):2519-27. doi: 10.1007/s00424-015-1728-y. Epub 2015 Aug 26. Pflugers Arch. 2015. PMID: 26306541
-
Unique cysteine-enriched, D2L5 and D4L6 extracellular loops in CaV3 T-type channels alter the passage and block of monovalent and divalent ions.Sci Rep. 2020 Jul 24;10(1):12404. doi: 10.1038/s41598-020-69197-3. Sci Rep. 2020. PMID: 32710088 Free PMC article.
-
Cav3 T-type channels: regulators for gating, membrane expression, and cation selectivity.Pflugers Arch. 2014 Apr;466(4):645-60. doi: 10.1007/s00424-014-1449-7. Epub 2014 Feb 11. Pflugers Arch. 2014. PMID: 24515291 Review.
-
A mutually induced conformational fit underlies Ca2+-directed interactions between calmodulin and the proximal C terminus of KCNQ4 K+ channels.J Biol Chem. 2019 Apr 12;294(15):6094-6112. doi: 10.1074/jbc.RA118.006857. Epub 2019 Feb 26. J Biol Chem. 2019. PMID: 30808708 Free PMC article.
Cited by
-
A governance of ion selectivity based on the occupancy of the "beacon" in one- and four-domain calcium and sodium channels.Channels (Austin). 2023 Dec;17(1):2191773. doi: 10.1080/19336950.2023.2191773. Channels (Austin). 2023. PMID: 37075164 Free PMC article.
-
Calenduloside E suppresses calcium overload by promoting the interaction between L-type calcium channels and Bcl2-associated athanogene 3 to alleviate myocardial ischemia/reperfusion injury.J Adv Res. 2020 Oct 31;34:173-186. doi: 10.1016/j.jare.2020.10.005. eCollection 2021 Dec. J Adv Res. 2020. PMID: 35024189 Free PMC article.
-
Developmentally Regulated Rebound Depolarization Enhances Spike Timing Precision in Auditory Midbrain Neurons.Front Cell Neurosci. 2020 Aug 6;14:236. doi: 10.3389/fncel.2020.00236. eCollection 2020. Front Cell Neurosci. 2020. PMID: 32848625 Free PMC article.
-
The mechanism of complex formation between calmodulin and voltage gated calcium channels revealed by molecular dynamics.PLoS One. 2021 Oct 5;16(10):e0258112. doi: 10.1371/journal.pone.0258112. eCollection 2021. PLoS One. 2021. PMID: 34610038 Free PMC article.
-
Neuronal Cav3 channelopathies: recent progress and perspectives.Pflugers Arch. 2020 Jul;472(7):831-844. doi: 10.1007/s00424-020-02429-7. Epub 2020 Jul 7. Pflugers Arch. 2020. PMID: 32638069 Free PMC article. Review.
References
-
- Ben-Johny M., Dick I. E., Sang L., Limpitikul W. B., Kang P. W., Niu J., Banerjee R., Yang W., Babich J. S., Issa J. B., Lee S. R., Namkung H., Li J., Zhang M., Yang P. S., et al. (2015) Towards a unified theory of calmodulin regulation (calmodulation) of voltage-gated calcium and sodium channels. Curr. Mol. Pharmacol. 8, 188–205 - PMC - PubMed
-
- Budde T., Meuth S., and Pape H. C. (2002) Calcium-dependent inactivation of neuronal calcium channels. Nat. Rev. Neurosci. 3, 873–883 - PubMed
-
- Halling D. B., Aracena-Parks P., and Hamilton S. L. (2006) Regulation of voltage-gated Ca2+ channels by calmodulin. Sci. STKE 2006, er1. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
