

Hydrogen Gas Is Involved in Auxin-Induced Lateral Root Formation by Modulating Nitric Oxide Synthesis
- PMID: 28972563
- PMCID: PMC5666766
- DOI: 10.3390/ijms18102084
Hydrogen Gas Is Involved in Auxin-Induced Lateral Root Formation by Modulating Nitric Oxide Synthesis
Abstract
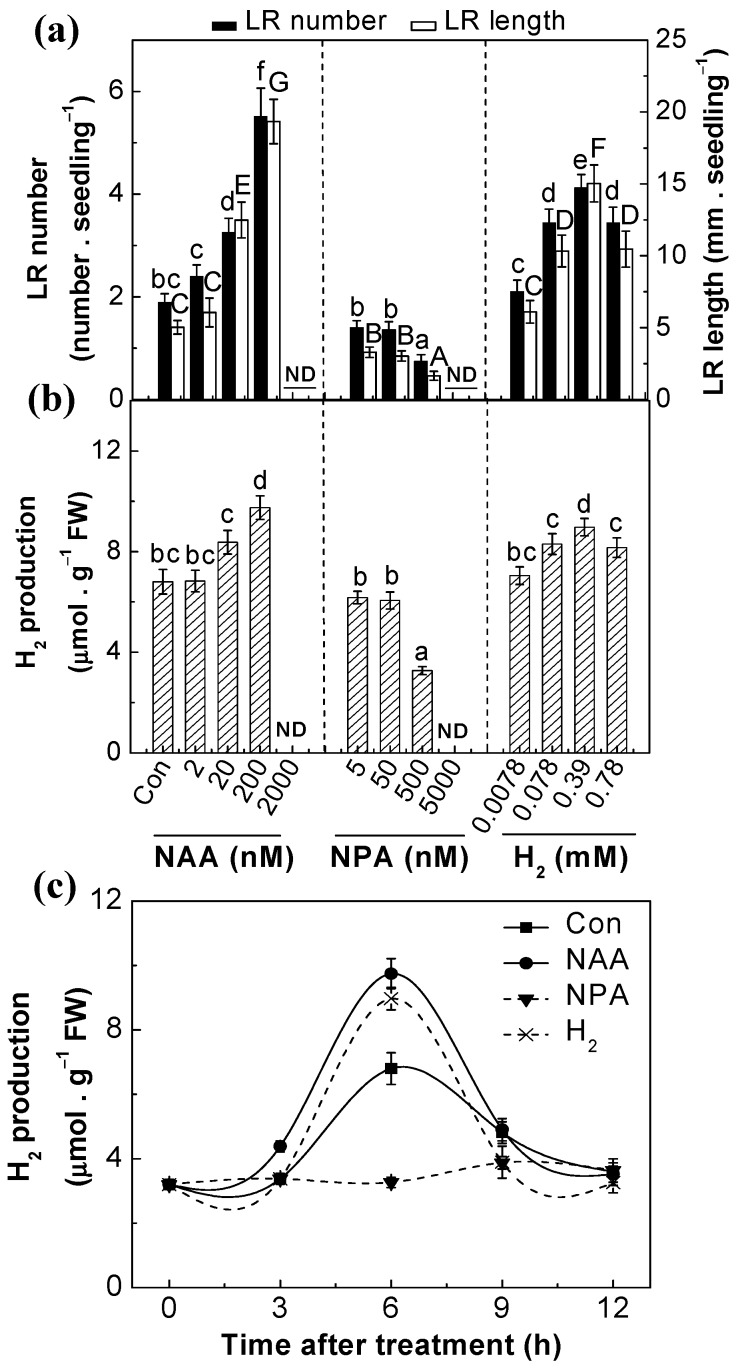
Metabolism of molecular hydrogen (H₂) in bacteria and algae has been widely studied, and it has attracted increasing attention in the context of animals and plants. However, the role of endogenous H₂ in lateral root (LR) formation is still unclear. Here, our results showed that H₂-induced lateral root formation is a universal event. Naphthalene-1-acetic acid (NAA; the auxin analog) was able to trigger endogenous H₂ production in tomato seedlings, and a contrasting response was observed in the presence of N-1-naphthyphthalamic acid (NPA), an auxin transport inhibitor. NPA-triggered the inhibition of H₂ production and thereafter lateral root development was rescued by exogenously applied H₂. Detection of endogenous nitric oxide (NO) by the specific probe 4-amino-5-methylamino-2',7'-difluorofluorescein diacetate (DAF-FM DA) and electron paramagnetic resonance (EPR) analyses revealed that the NO level was increased in both NAA- and H₂-treated tomato seedlings. Furthermore, NO production and thereafter LR formation induced by auxin and H₂ were prevented by 2-4-carboxyphenyl-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (cPTIO; a specific scavenger of NO) and the inhibitor of nitrate reductase (NR; an important NO synthetic enzyme). Molecular evidence confirmed that some representative NO-targeted cell cycle regulatory genes were also induced by H₂, but was impaired by the removal of endogenous NO. Genetic evidence suggested that in the presence of H₂, Arabidopsis mutants nia2 (in particular) and nia1 (two nitrate reductases (NR)-defective mutants) exhibited defects in lateral root length. Together, these results demonstrated that auxin-induced H₂ production was associated with lateral root formation, at least partially via a NR-dependent NO synthesis.
Keywords: H2; NO; auxin; lateral root formation; nitrate reductase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Perrine-Walker F.M., Jublanc E. The localization of auxin transporters PIN3 and LAX3 during lateral root development in Arabidopsis thaliana. Biol. Plantarum. 2014;58:778–782. doi: 10.1007/s10535-014-0446-1. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
