

The evolutionary origins of Lévy walk foraging
- PMID: 28972973
- PMCID: PMC5640246
- DOI: 10.1371/journal.pcbi.1005774
The evolutionary origins of Lévy walk foraging
Abstract
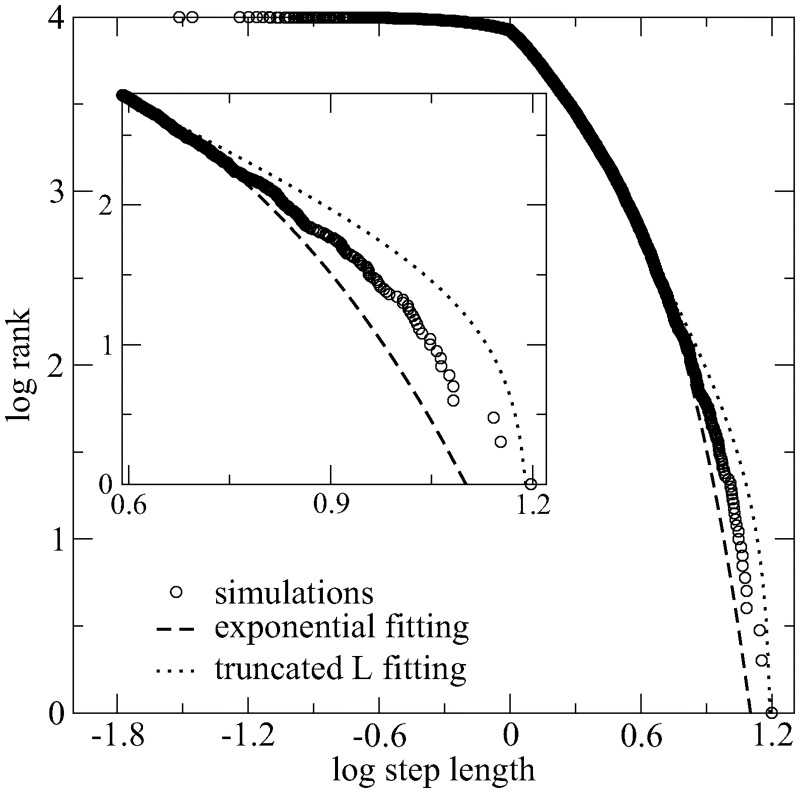
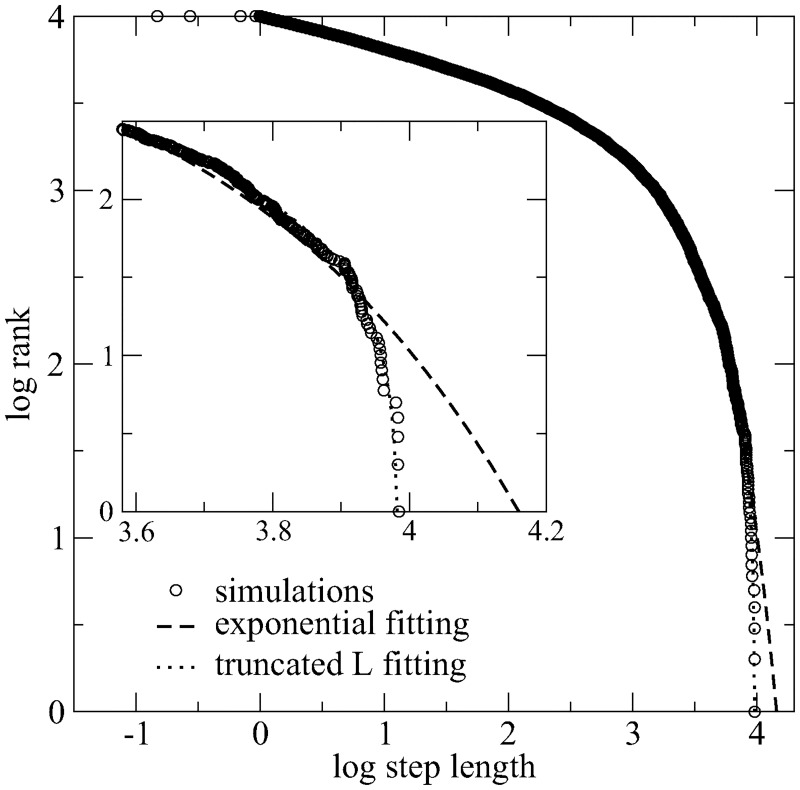
We study through a reaction-diffusion algorithm the influence of landscape diversity on the efficiency of search dynamics. Remarkably, the identical optimal search strategy arises in a wide variety of environments, provided the target density is sparse and the searcher's information is restricted to its close vicinity. Our results strongly impact the current debate on the emergentist vs. evolutionary origins of animal foraging. The inherent character of the optimal solution (i.e., independent on the landscape for the broad scenarios assumed here) suggests an interpretation favoring the evolutionary view, as originally implied by the Lévy flight foraging hypothesis. The latter states that, under conditions of scarcity of information and sparse resources, some organisms must have evolved to exploit optimal strategies characterized by heavy-tailed truncated power-law distributions of move lengths. These results strongly suggest that Lévy strategies-and hence the selection pressure for the relevant adaptations-are robust with respect to large changes in habitat. In contrast, the usual emergentist explanation seems not able to explain how very similar Lévy walks can emerge from all the distinct non-Lévy foraging strategies that are needed for the observed large variety of specific environments. We also report that deviations from Lévy can take place in plentiful ecosystems, where locomotion truncation is very frequent due to high encounter rates. So, in this case normal diffusion strategies-performing as effectively as the optimal one-can naturally emerge from Lévy. Our results constitute the strongest theoretical evidence to date supporting the evolutionary origins of experimentally observed Lévy walks.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
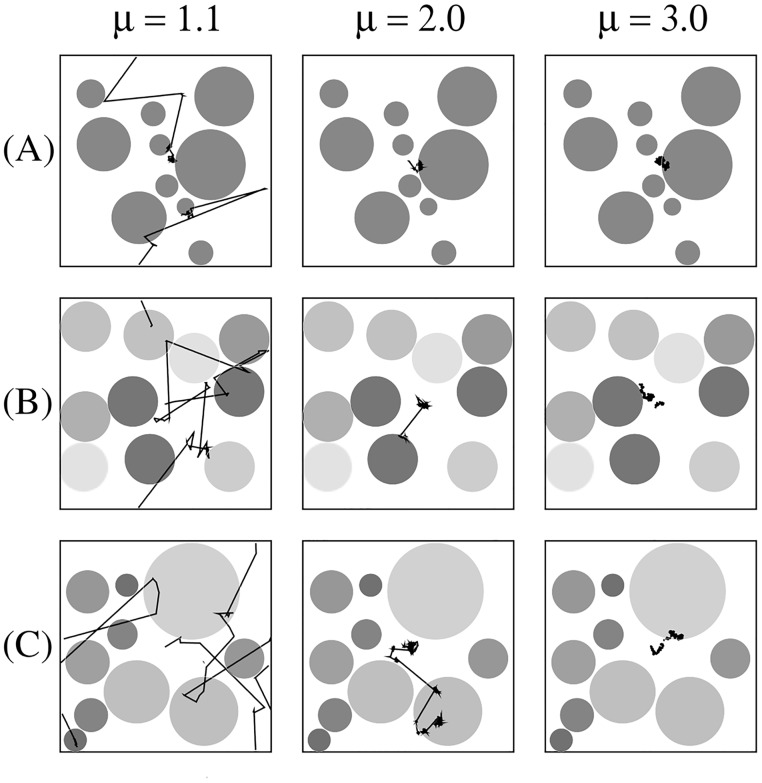
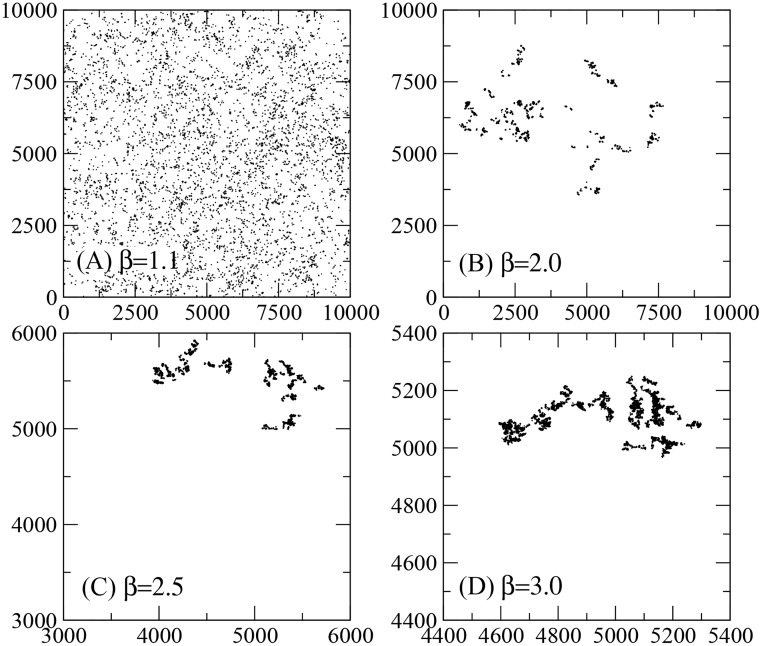
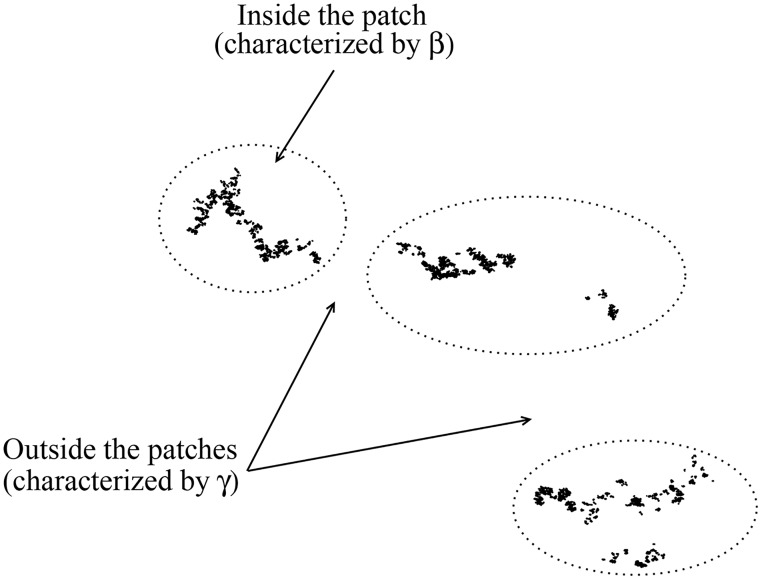
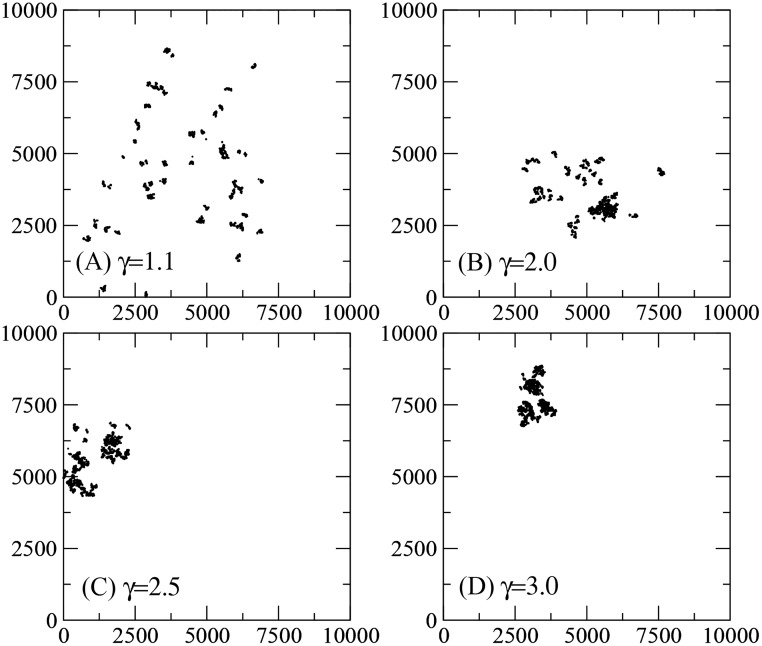
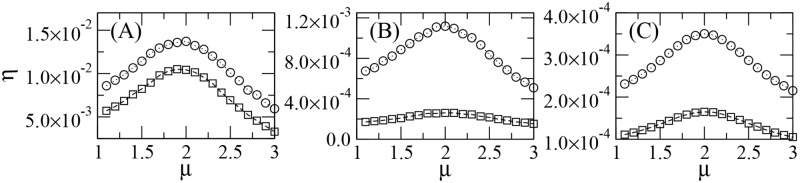
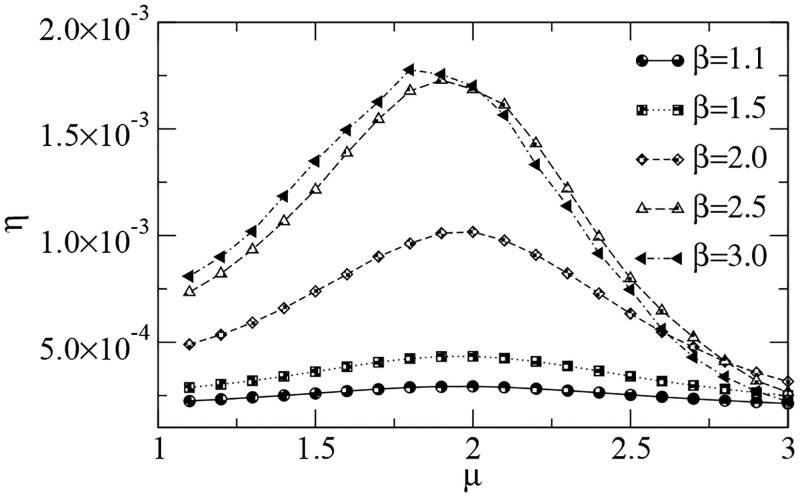
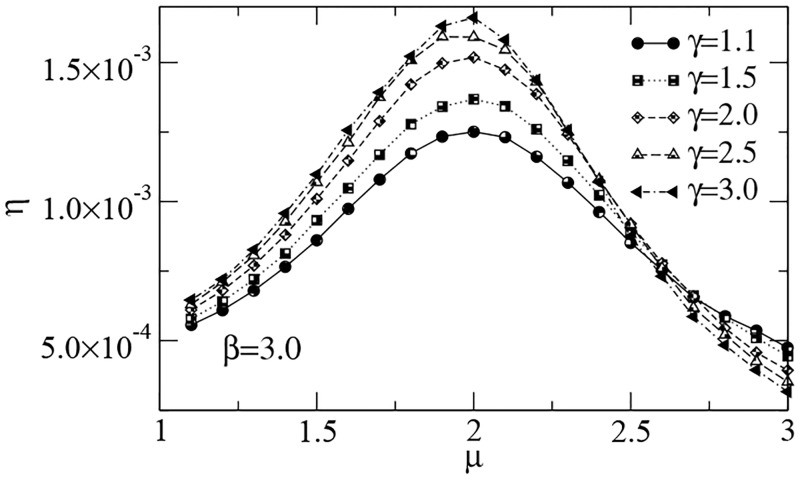
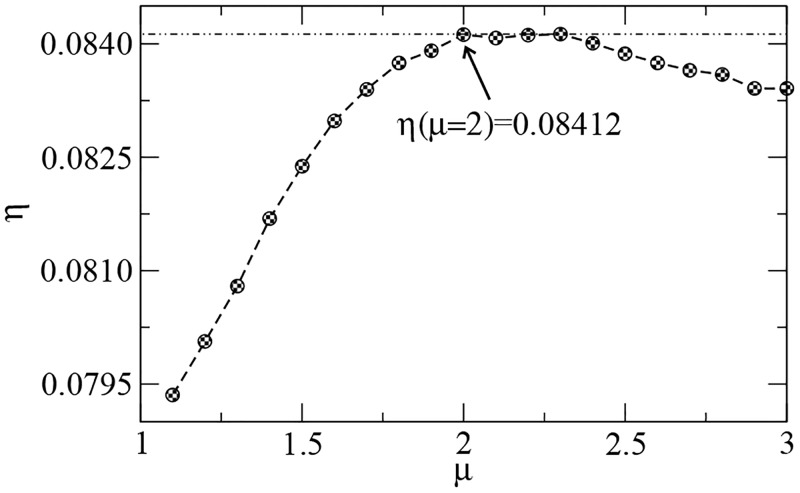
References
-
- Viswanathan GM, da Luz MGE, Raposo EP, Stanley HE. The Physics of Foraging: An Introduction to Random Searches and Biological Encounters. Cambridge: Cambridge University Press; 2011.
-
- Méndez V, Campos D, Bartumeus F. Stochastic Foundations in Movement Ecology: Anomalous Diffusion, Front Propagation and Random Searches. Berlin: Springer; 2014.
-
- MacArthur RH, Pianka ER. On optimal use of a patchy environment. Amer Natur. 1966. November;100:603–9. 10.1086/282454 - DOI
-
- Pyke GH. Optimal Foraging Theory: A Critical Review. Annu Rev Ecol Syst. 1984;15:523–75. 10.1146/annurev.es.15.110184.002515 - DOI
-
- Kamil AC, Sargent TD (eds.) Foraging Behavior: Ecological, Ethnological, and Psychological Approaches. New York: Garland STPM Press; 1981.
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
