

Transcription-induced supercoiling explains formation of self-interacting chromatin domains in S. pombe
- PMID: 28973473
- PMCID: PMC5622301
- DOI: 10.1093/nar/gkx716
Transcription-induced supercoiling explains formation of self-interacting chromatin domains in S. pombe
Abstract
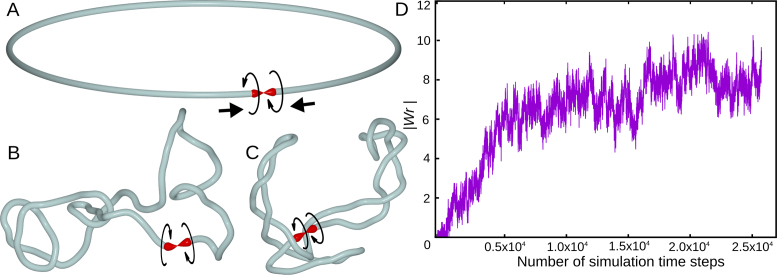
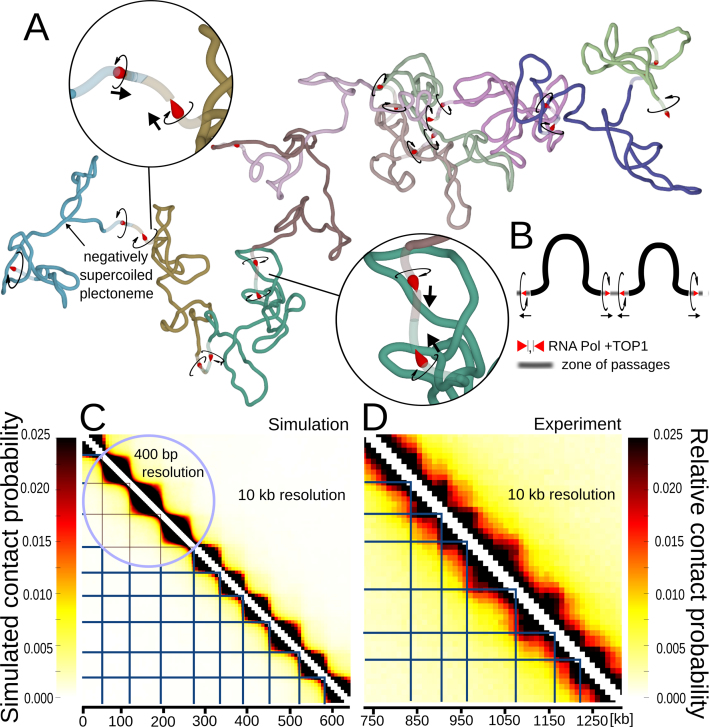
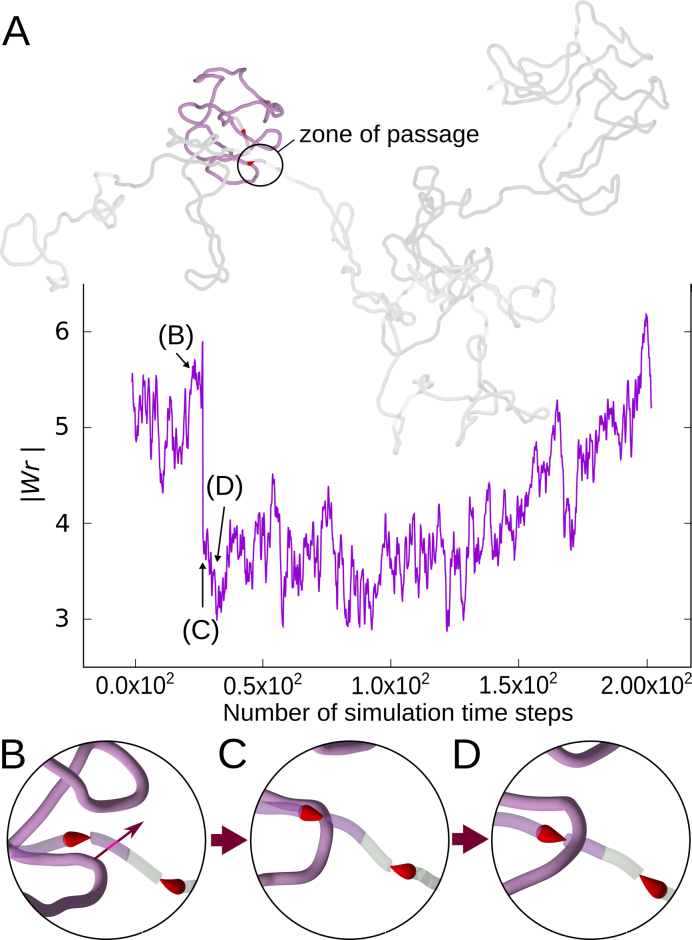
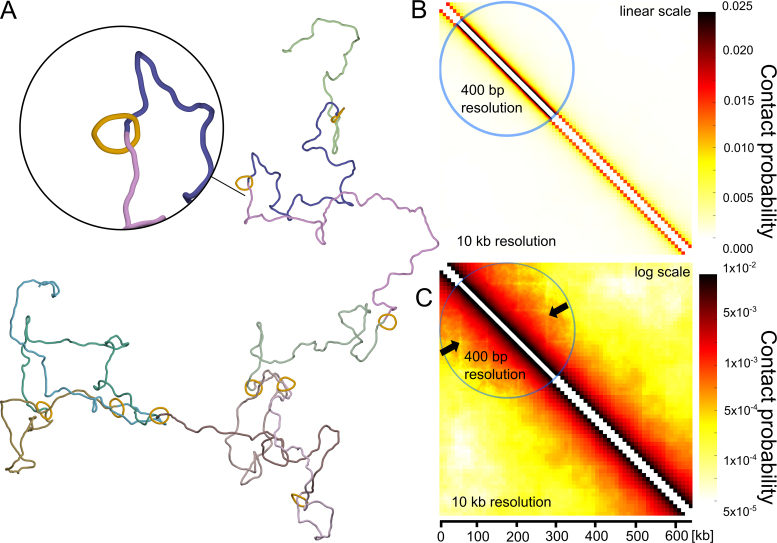
The question of how self-interacting chromatin domains in interphase chromosomes are structured and generated dominates current discussions on eukaryotic chromosomes. Numerical simulations using standard polymer models have been helpful in testing the validity of various models of chromosome organization. Experimental contact maps can be compared with simulated contact maps and thus verify how good is the model. With increasing resolution of experimental contact maps, it became apparent though that active processes need to be introduced into models to recapitulate the experimental data. Since transcribing RNA polymerases are very strong molecular motors that induce axial rotation of transcribed DNA, we present here models that include such rotational motors. We also include into our models swivels and sites for intersegmental passages that account for action of DNA topoisomerases releasing torsional stress. Using these elements in our models, we show that transcription-induced supercoiling generated in the regions with divergent-transcription and supercoiling relaxation occurring between these regions are sufficient to explain formation of self-interacting chromatin domains in chromosomes of fission yeast (S. pombe).
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Sexton T., Yaffe E., Kenigsberg E., Bantignies F., Leblanc B., Hoichman M., Parrinello H., Tanay A., Cavalli G.. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell. 2012; 148:458–472. - PubMed
-
- Sanborn A.L., Rao S.S., Huang S.C., Durand N.C., Huntley M.H., Jewett A.I., Bochkov I.D., Chinnappan D., Cutkosky A., Li J. et al. . Chromatin extrusion explains key features of loop and domain formation in wild-type and engineered genomes. Proc. Natl. Acad. Sci. U.S.A. 2015; 112:E6456–E6465. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
