

Nucleus accumbens feedforward inhibition circuit promotes cocaine self-administration
- PMID: 28973852
- PMCID: PMC5642706
- DOI: 10.1073/pnas.1707822114
Nucleus accumbens feedforward inhibition circuit promotes cocaine self-administration
Abstract
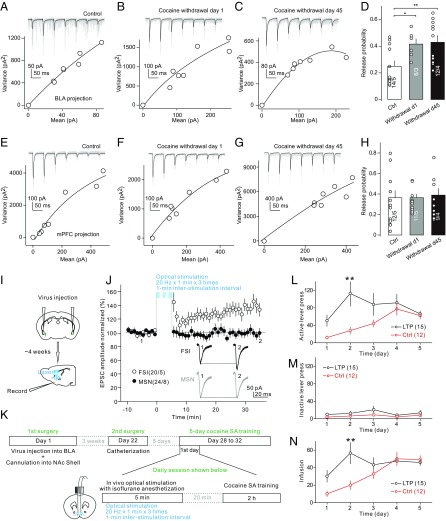
The basolateral amygdala (BLA) sends excitatory projections to the nucleus accumbens (NAc) and regulates motivated behaviors partially by activating NAc medium spiny neurons (MSNs). Here, we characterized a feedforward inhibition circuit, through which BLA-evoked activation of NAc shell (NAcSh) MSNs was fine-tuned by GABAergic monosynaptic innervation from adjacent fast-spiking interneurons (FSIs). Specifically, BLA-to-NAcSh projections predominantly innervated NAcSh FSIs compared with MSNs and triggered action potentials in FSIs preceding BLA-mediated activation of MSNs. Due to these anatomical and temporal properties, activation of the BLA-to-NAcSh projection resulted in a rapid FSI-mediated inhibition of MSNs, timing-contingently dictating BLA-evoked activation of MSNs. Cocaine self-administration selectively and persistently up-regulated the presynaptic release probability of BLA-to-FSI synapses, entailing enhanced FSI-mediated feedforward inhibition of MSNs upon BLA activation. Experimentally enhancing the BLA-to-FSI transmission in vivo expedited the acquisition of cocaine self-administration. These results reveal a previously unidentified role of an FSI-embedded circuit in regulating NAc-based drug seeking and taking.
Keywords: basolateral amygdala; cocaine addiction; fast-spiking interneuron; medium spiny neuron; synaptic plasticity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
-
- Pennartz CM, Groenewegen HJ, Lopes da Silva FH. The nucleus accumbens as a complex of functionally distinct neuronal ensembles: An integration of behavioural, electrophysiological and anatomical data. Prog Neurobiol. 1994;42:719–761. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
