

Universal poroelastic mechanism for hydraulic signals in biomimetic and natural branches
- PMID: 28973910
- PMCID: PMC5651757
- DOI: 10.1073/pnas.1707675114
Universal poroelastic mechanism for hydraulic signals in biomimetic and natural branches
Erratum in
-
Correction for Louf et al., Universal poroelastic mechanism for hydraulic signals in biomimetic and natural branches.Proc Natl Acad Sci U S A. 2017 Oct 31;114(44):E9423. doi: 10.1073/pnas.1717494114. Epub 2017 Oct 23. Proc Natl Acad Sci U S A. 2017. PMID: 29078426 Free PMC article. No abstract available.
Abstract
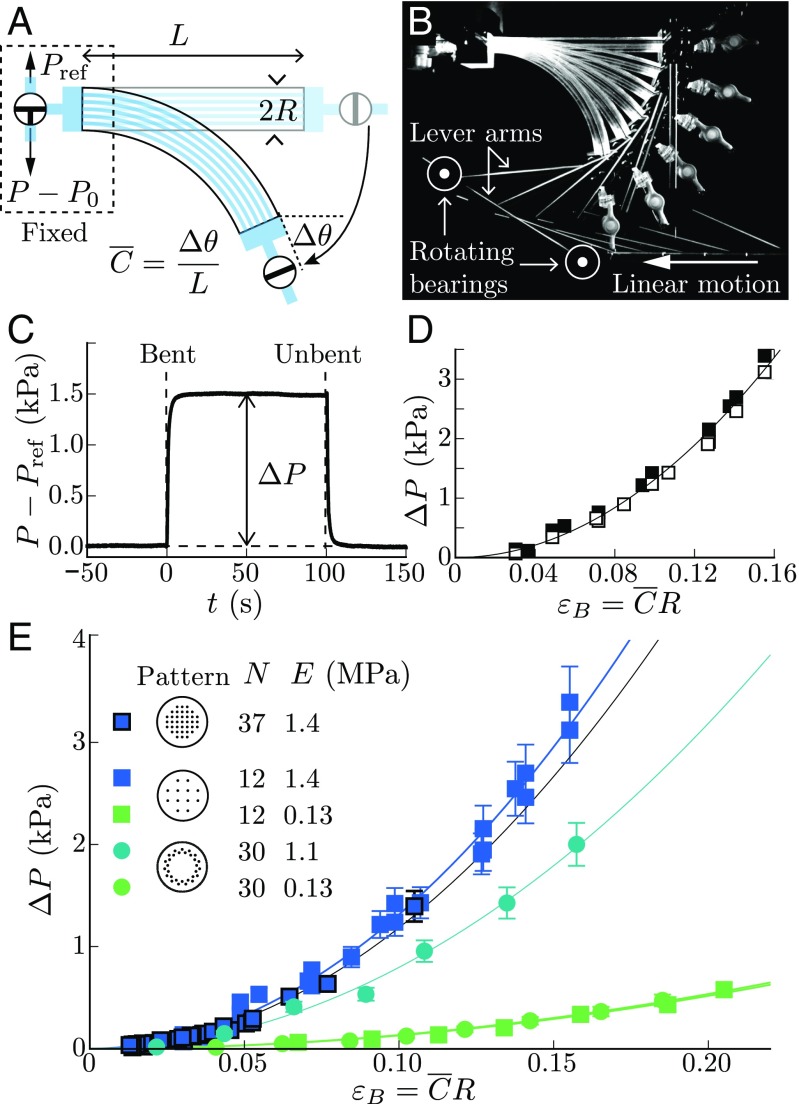
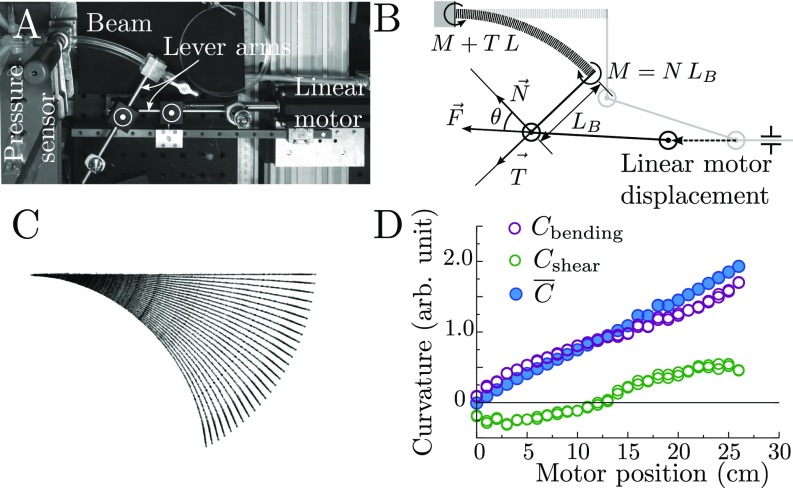
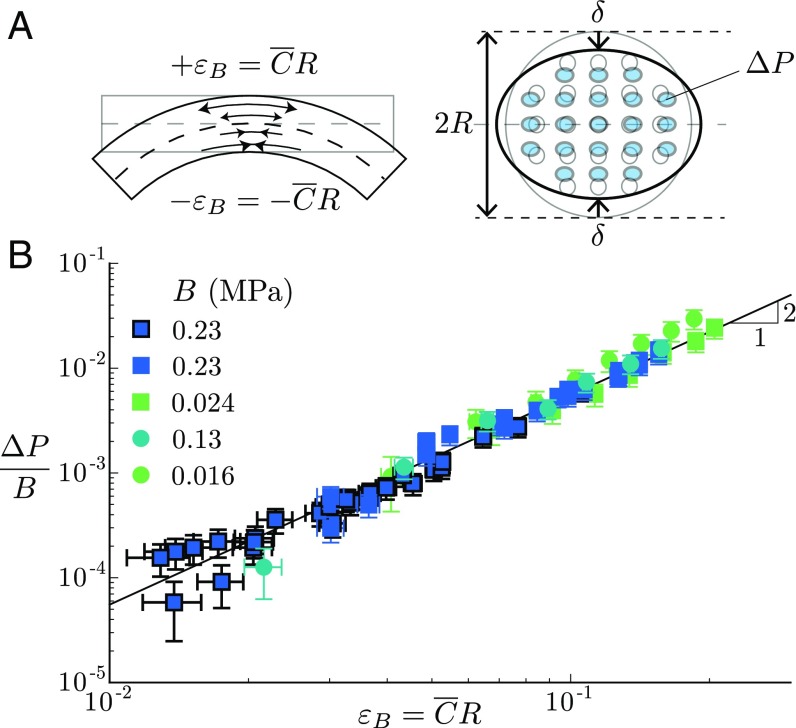
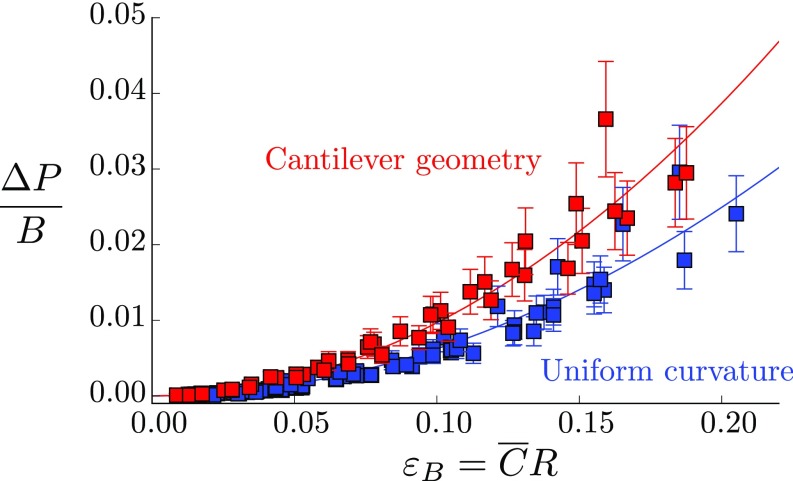
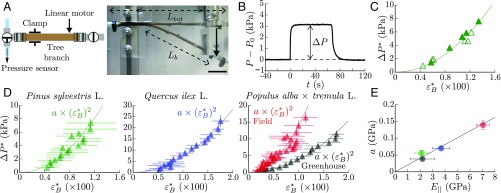
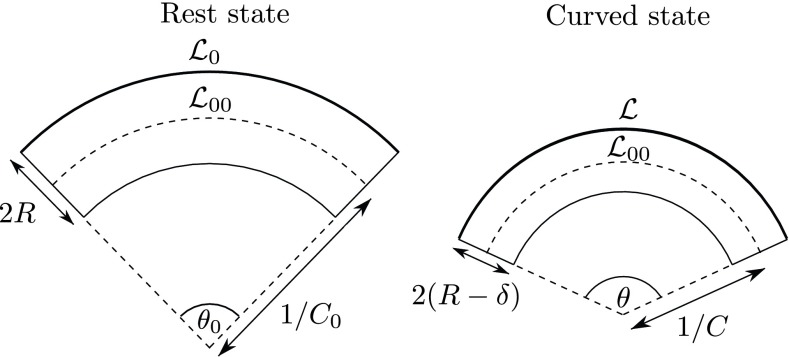
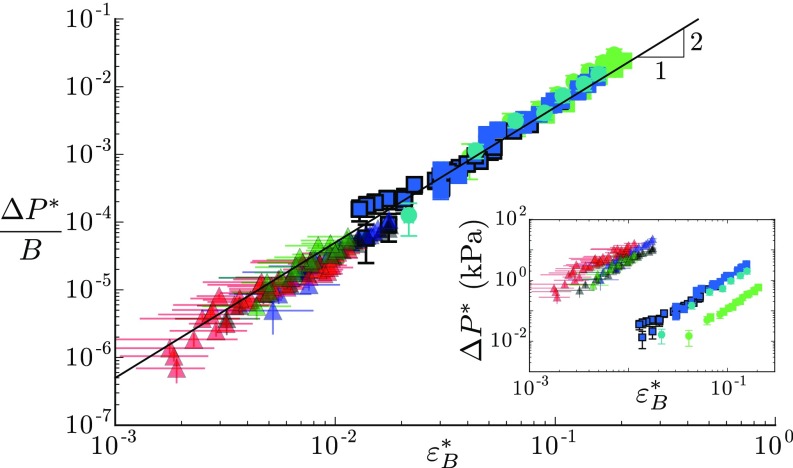
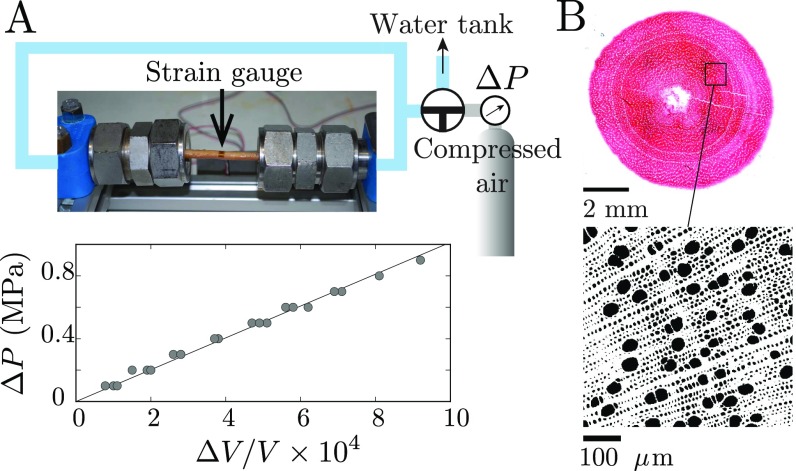
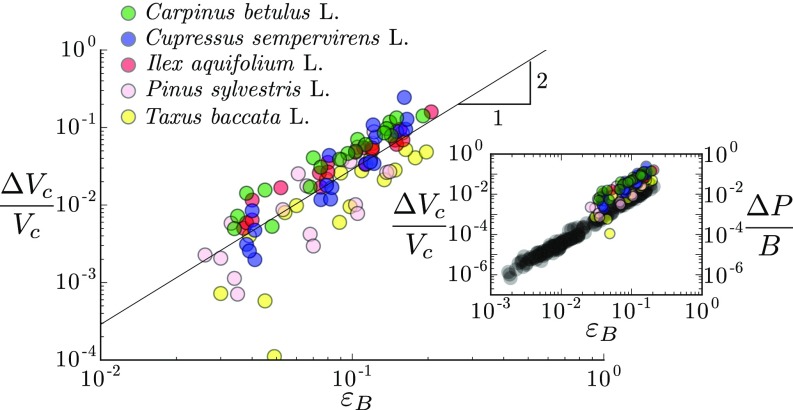
Plants constantly undergo external mechanical loads such as wind or touch and respond to these stimuli by acclimating their growth processes. A fascinating feature of this mechanical-induced growth response is that it can occur rapidly and at long distance from the initial site of stimulation, suggesting the existence of a fast signal that propagates across the whole plant. The nature and origin of the signal is still not understood, but it has been recently suggested that it could be purely mechanical and originate from the coupling between the local deformation of the tissues (bending) and the water pressure in the plant vascular system. Here, we address the physical origin of this hydromechanical coupling using a biomimetic strategy. We designed soft artificial branches perforated with longitudinal liquid-filled channels that mimic the basic features of natural stems and branches. In response to bending, a strong overpressure is generated in the channels that varies quadratically with the bending curvature. A model based on a mechanism analogous to the ovalization of hollow tubes enables us to predict quantitatively this nonlinear poroelastic response and identify the key physical parameters that control the generation of the pressure pulse. Further experiments conducted on natural tree branches reveal the same phenomenology. Once rescaled by the model prediction, both the biomimetic and natural branches fall on the same master curve, enlightening the universality of our poroelastic mechanism for the generation of hydraulic signals in plants.
Keywords: biomimetism; long-distance signaling; nonlinear beams; plant biomechanics; poroelasticity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Darwin C, Darwin F. The Power of Movement in Plants. Murray; London: 1880.
-
- Knight TA. Account of some experiments on the descent of the sap in trees. Phil Trans Roy Soc Lond. 1803;93:277–289.
-
- Jaffe MJ, Leopold AC, Staples RC. Thigmo responses in plants and fungi. Am J Bot. 2002;89:375–382. - PubMed
-
- Braam J. In touch: Plant responses to mechanical stimuli. New Phytol. 2005;165:373–389. - PubMed
-
- Telewski FW. A unified hypothesis of mechanoperception in plants. Am J Bot. 2006;93:1466–1476. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
