

Discovery of an O-mannosylation pathway selectively serving cadherins and protocadherins
- PMID: 28973932
- PMCID: PMC5651762
- DOI: 10.1073/pnas.1708319114
Discovery of an O-mannosylation pathway selectively serving cadherins and protocadherins
Abstract
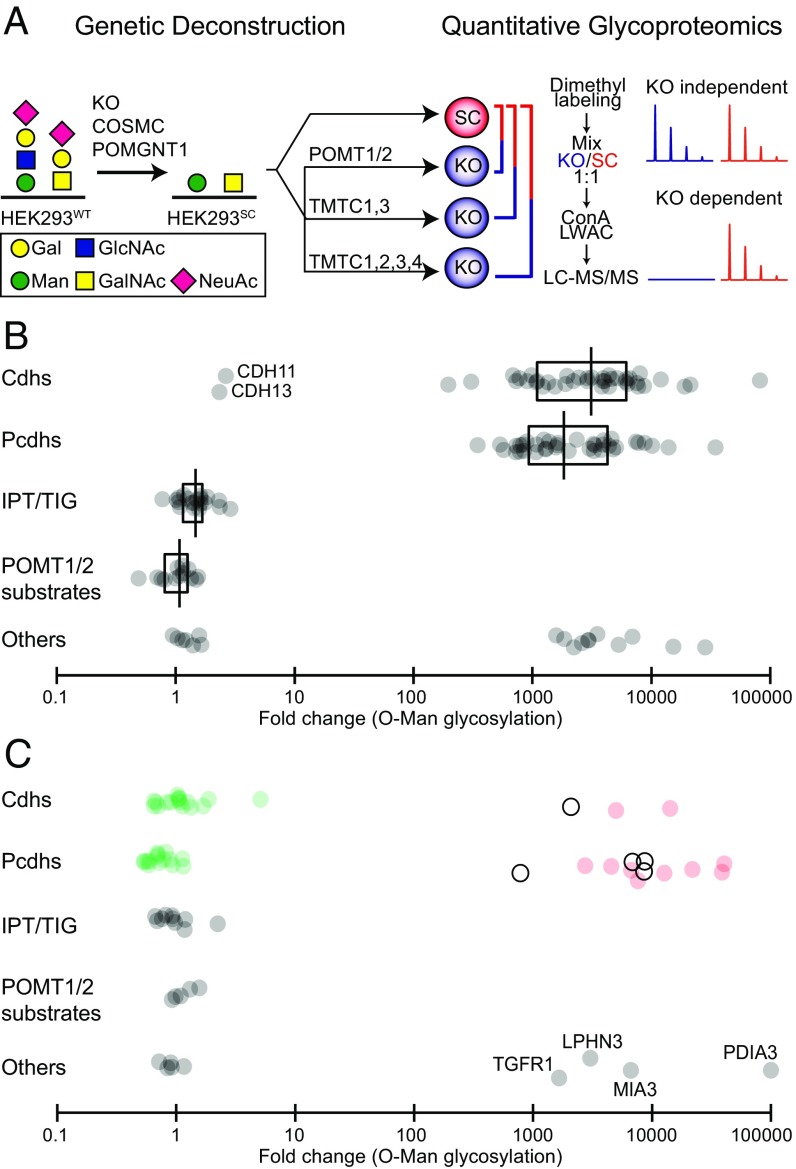
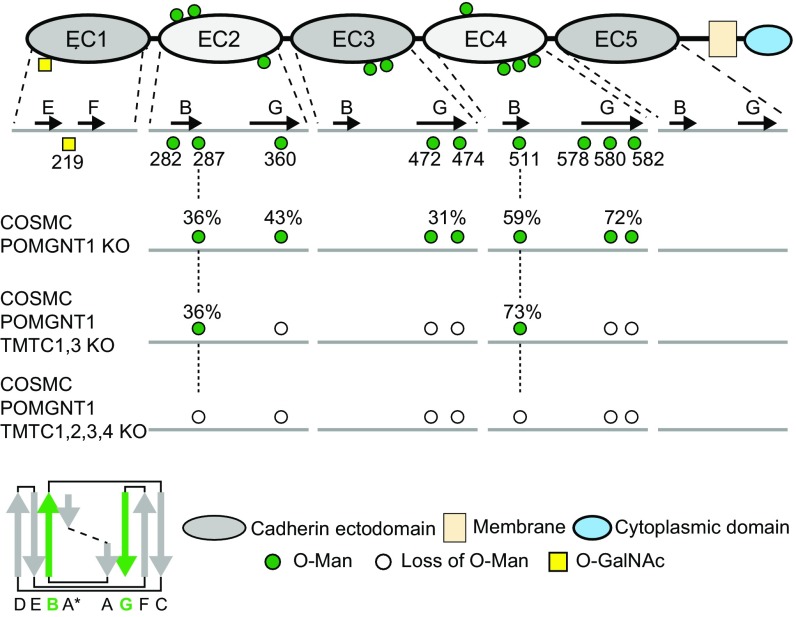
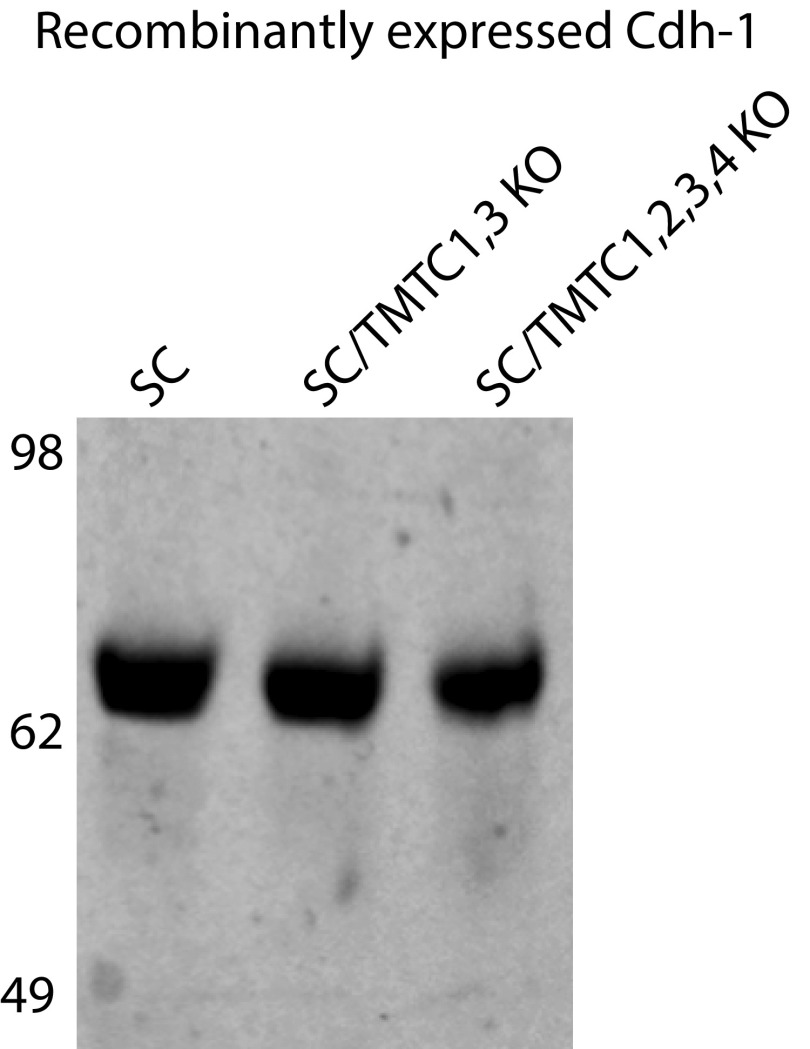
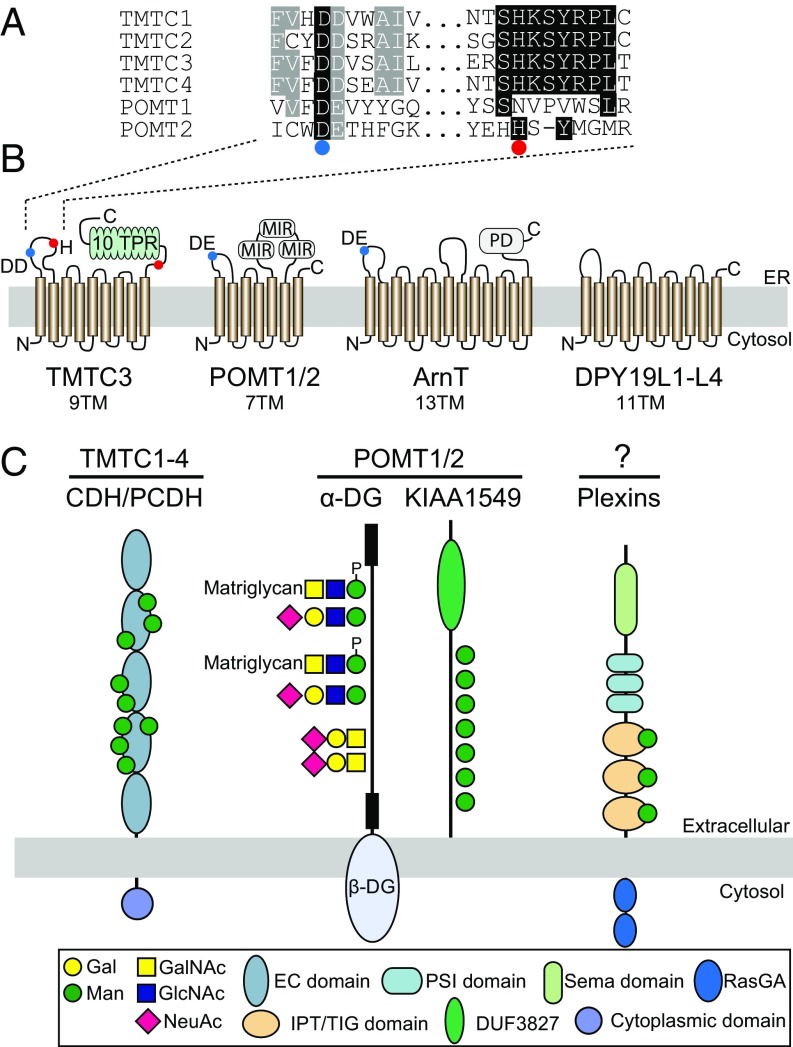
The cadherin (cdh) superfamily of adhesion molecules carry O-linked mannose (O-Man) glycans at highly conserved sites localized to specific β-strands of their extracellular cdh (EC) domains. These O-Man glycans do not appear to be elongated like O-Man glycans found on α-dystroglycan (α-DG), and we recently demonstrated that initiation of cdh/protocadherin (pcdh) O-Man glycosylation is not dependent on the evolutionary conserved POMT1/POMT2 enzymes that initiate O-Man glycosylation on α-DG. Here, we used a CRISPR/Cas9 genetic dissection strategy combined with sensitive and quantitative O-Man glycoproteomics to identify a homologous family of four putative protein O-mannosyltransferases encoded by the TMTC1-4 genes, which were found to be imperative for cdh and pcdh O-Man glycosylation. KO of all four TMTC genes in HEK293 cells resulted in specific loss of cdh and pcdh O-Man glycosylation, whereas combined KO of TMTC1 and TMTC3 resulted in selective loss of O-Man glycans on specific β-strands of EC domains, suggesting that each isoenzyme serves a different function. In addition, O-Man glycosylation of IPT/TIG domains of plexins and hepatocyte growth factor receptor was not affected in TMTC KO cells, suggesting the existence of yet another O-Man glycosylation machinery. Our study demonstrates that regulation of O-mannosylation in higher eukaryotes is more complex than envisioned, and the discovery of the functions of TMTCs provide insight into cobblestone lissencephaly caused by deficiency in TMTC3.
Keywords: O-glycosylation; gene editing; glycoproteomics; glycosyltransferase; mass spectrometry.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Lommel M, Strahl S. Protein O-mannosylation: Conserved from bacteria to humans. Glycobiology. 2009;19:816–828. - PubMed
-
- Loibl M, Strahl S. Protein O-mannosylation: What we have learned from baker’s yeast. Biochim Biophys Acta. 2013;1833:2438–2446. - PubMed
-
- Halim A, Anonsen JH. Microbial glycoproteomics. Curr Opin Struct Biol. 2017;44:143–150. - PubMed
-
- Barresi R, Campbell KP. Dystroglycan: From biosynthesis to pathogenesis of human disease. J Cell Sci. 2006;119:199–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
