

POLYGALACTURONASE INVOLVED IN EXPANSION3 Functions in Seedling Development, Rosette Growth, and Stomatal Dynamics in Arabidopsis thaliana
- PMID: 28974550
- PMCID: PMC5774581
- DOI: 10.1105/tpc.17.00568
POLYGALACTURONASE INVOLVED IN EXPANSION3 Functions in Seedling Development, Rosette Growth, and Stomatal Dynamics in Arabidopsis thaliana
Abstract
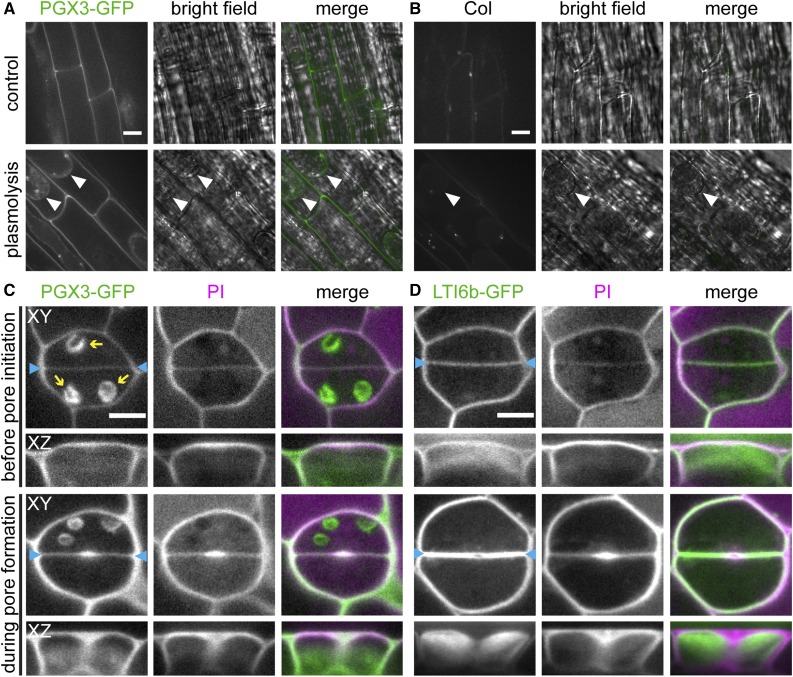
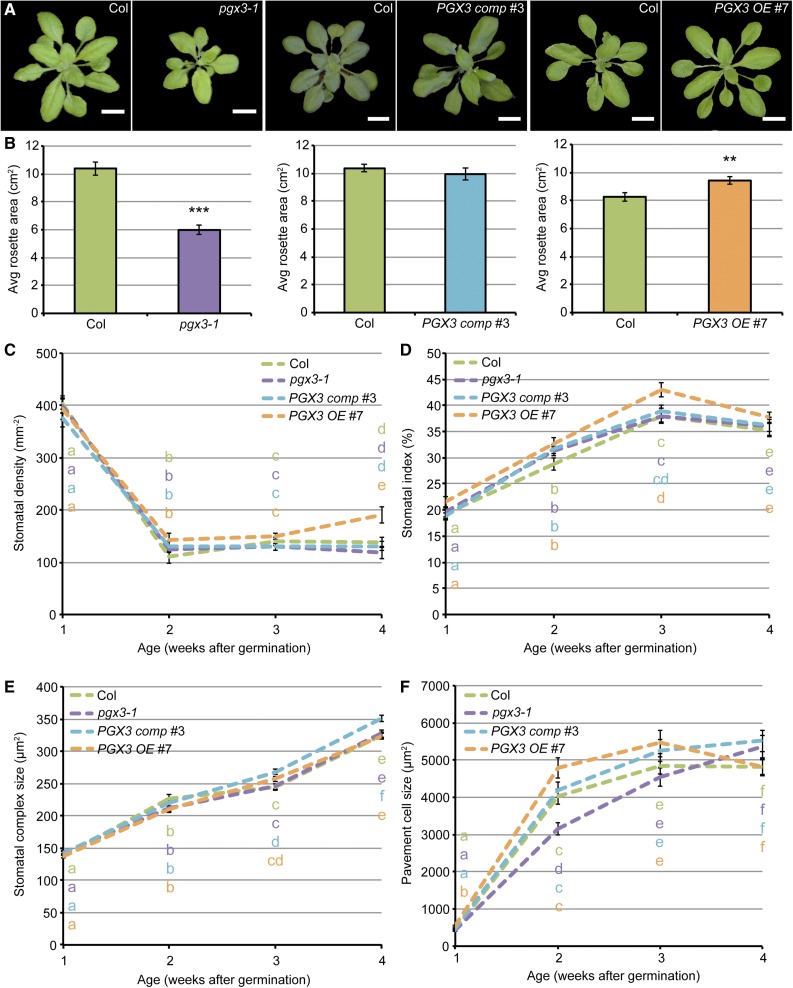
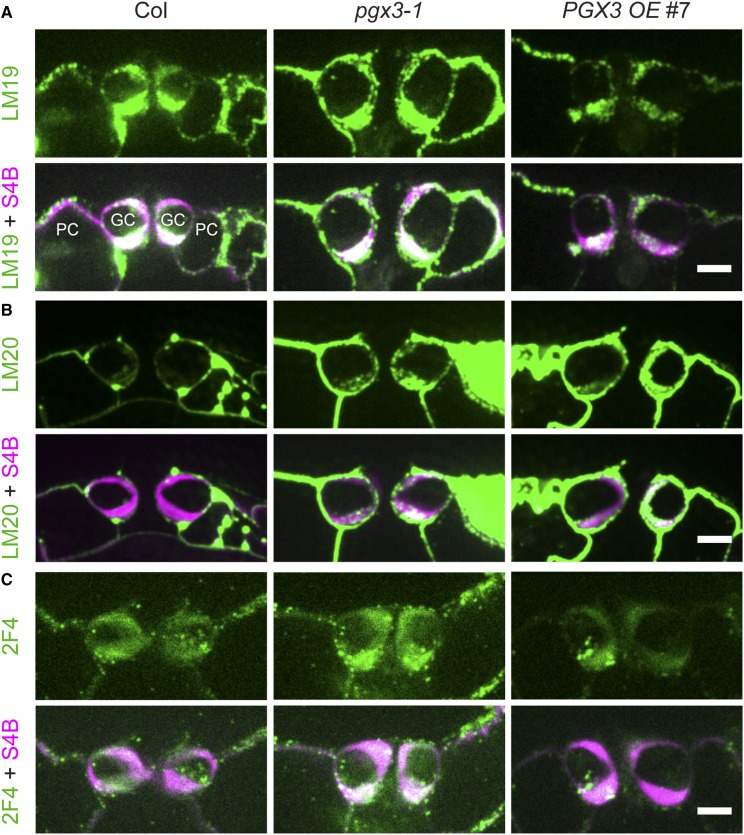
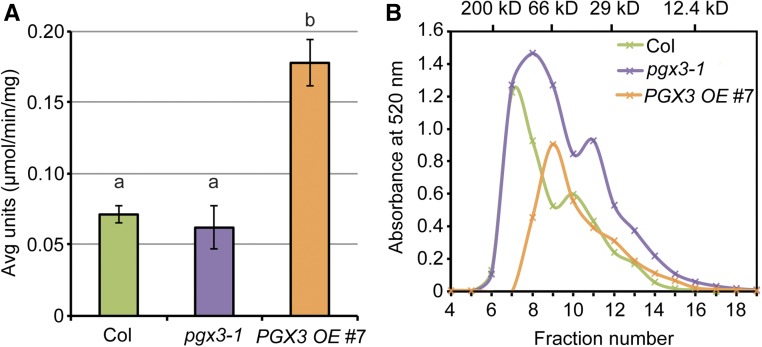
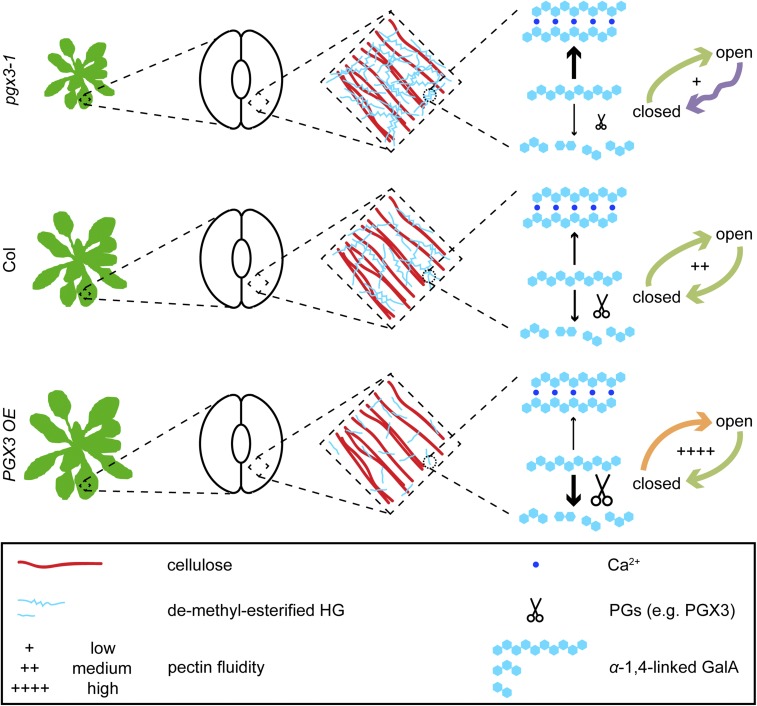
Plant cell separation and expansion require pectin degradation by endogenous pectinases such as polygalacturonases, few of which have been functionally characterized. Stomata are a unique system to study both processes because stomatal maturation involves limited separation between sister guard cells and stomatal responses require reversible guard cell elongation and contraction. However, the molecular mechanisms for how stomatal pores form and how guard cell walls facilitate dynamic stomatal responses remain poorly understood. We characterized POLYGALACTURONASE INVOLVED IN EXPANSION3 (PGX3), which is expressed in expanding tissues and guard cells. PGX3-GFP localizes to the cell wall and is enriched at sites of stomatal pore initiation in cotyledons. In seedlings, ablating or overexpressing PGX3 affects both cotyledon shape and the spacing and pore dimensions of developing stomata. In adult plants, PGX3 affects rosette size. Although stomata in true leaves display normal density and morphology when PGX3 expression is altered, loss of PGX3 prevents smooth stomatal closure, and overexpression of PGX3 accelerates stomatal opening. These phenotypes correspond with changes in pectin molecular mass and abundance that can affect wall mechanics. Together, these results demonstrate that PGX3-mediated pectin degradation affects stomatal development in cotyledons, promotes rosette expansion, and modulates guard cell mechanics in adult plants.
© 2017 American Society of Plant Biologists. All rights reserved.
Figures




References
-
- Atmodjo M.A., Hao Z., Mohnen D. (2013). Evolving views of pectin biosynthesis. Annu. Rev. Plant Biol. 64: 747–779. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
