

Differential interactions between Notch and ID factors control neurogenesis by modulating Hes factor autoregulation
- PMID: 28974640
- PMCID: PMC5665482
- DOI: 10.1242/dev.152520
Differential interactions between Notch and ID factors control neurogenesis by modulating Hes factor autoregulation
Abstract
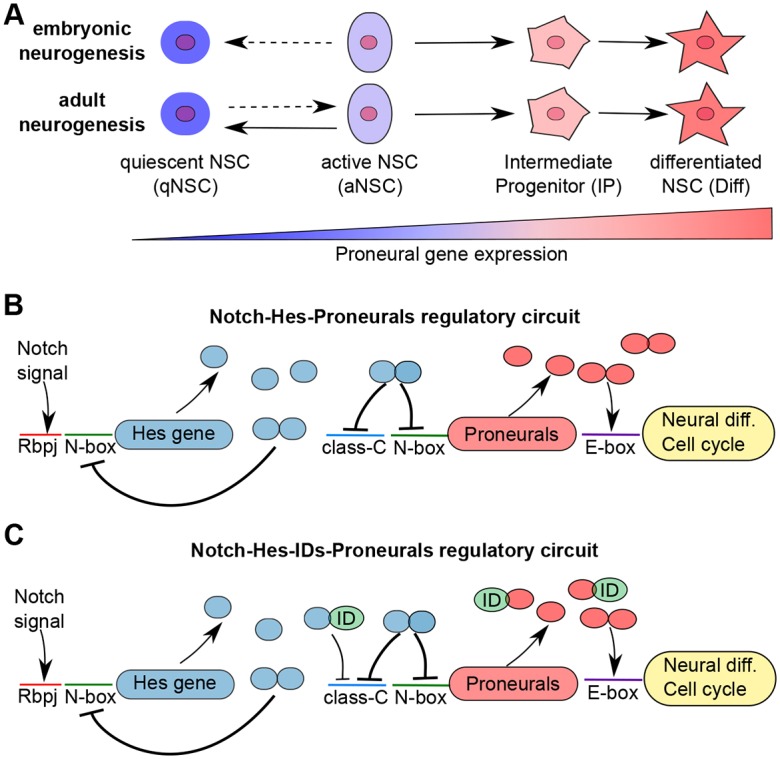
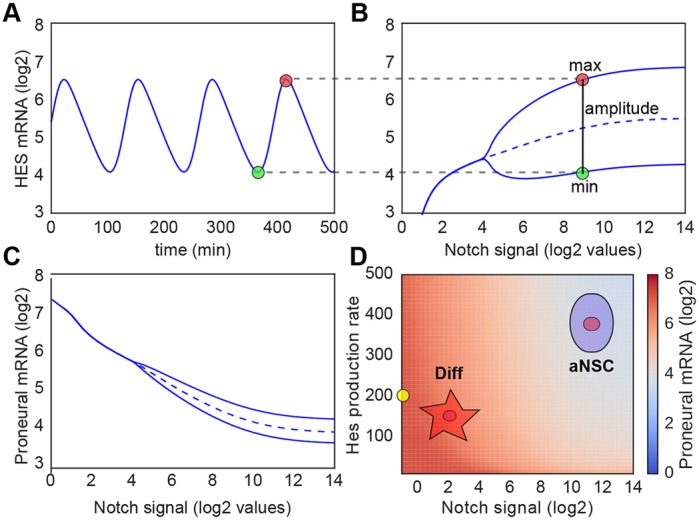
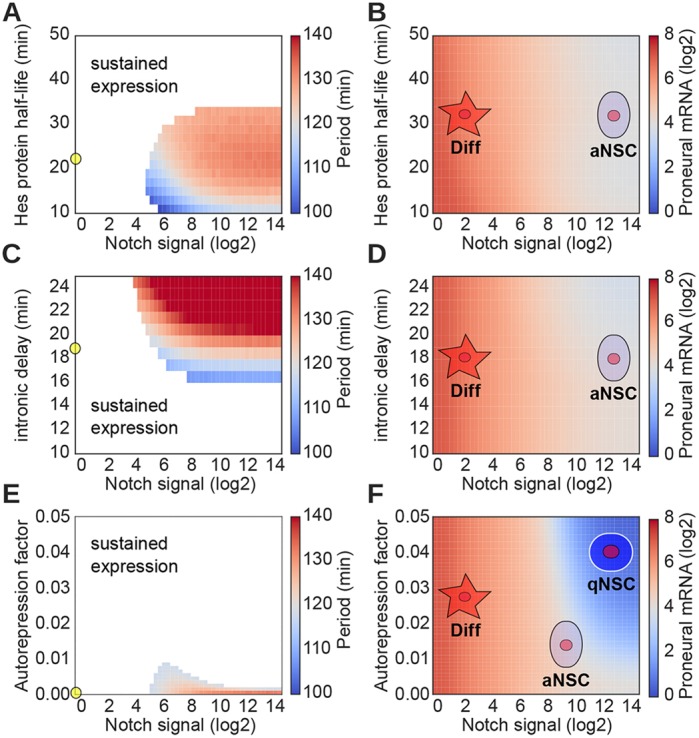
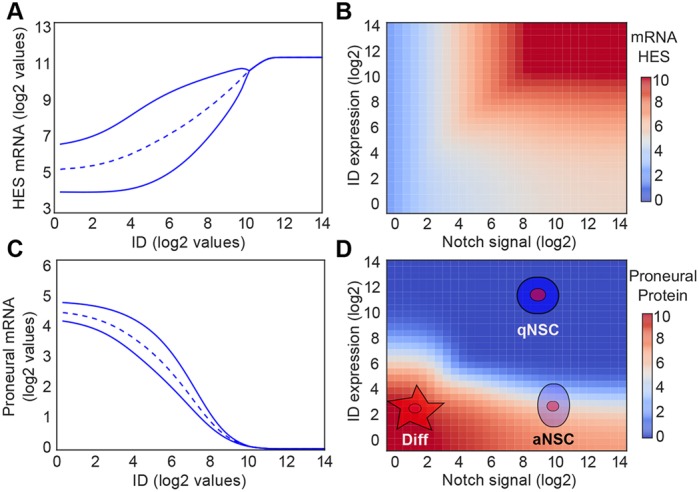
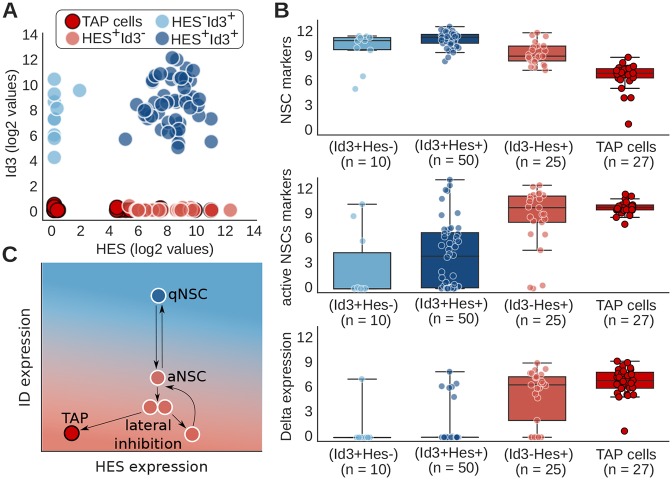
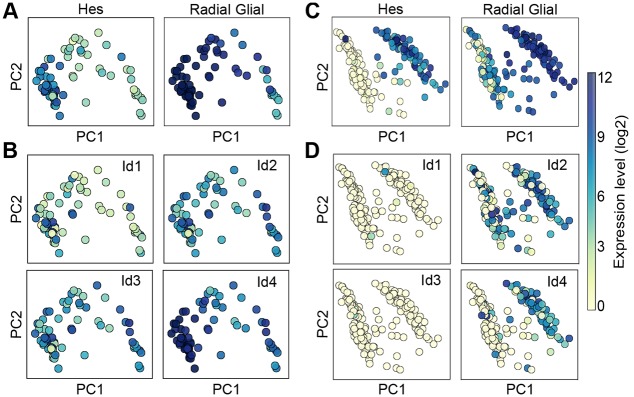
During embryonic and adult neurogenesis, neural stem cells (NSCs) generate the correct number and types of neurons in a temporospatial fashion. Control of NSC activity and fate is crucial for brain formation and homeostasis. Neurogenesis in the embryonic and adult brain differ considerably, but Notch signaling and inhibitor of DNA-binding (ID) factors are pivotal in both. Notch and ID factors regulate NSC maintenance; however, it has been difficult to evaluate how these pathways potentially interact. Here, we combined mathematical modeling with analysis of single-cell transcriptomic data to elucidate unforeseen interactions between the Notch and ID factor pathways. During brain development, Notch signaling dominates and directly regulates Id4 expression, preventing other ID factors from inducing NSC quiescence. Conversely, during adult neurogenesis, Notch signaling and Id2/3 regulate neurogenesis in a complementary manner and ID factors can induce NSC maintenance and quiescence in the absence of Notch. Our analyses unveil key molecular interactions underlying NSC maintenance and mechanistic differences between embryonic and adult neurogenesis. Similar Notch and ID factor interactions may be crucial in other stem cell systems.
Keywords: Computational biology; Id transcription factors; Neural stem cells; Neurogenesis; Notch signaling.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Andersen J., Urbán N., Achimastou A., Ito A., Simic M., Ullom K., Martynoga B., Lebel M., Göritz C., Frisén J. et al. (2014). A transcriptional mechanism integrating inputs from extracellular signals to activate hippocampal stem cells. Neuron 83, 1085-1097. 10.1016/j.neuron.2014.08.004 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
