

Molecular architecture of the PBP2-MreC core bacterial cell wall synthesis complex
- PMID: 28974686
- PMCID: PMC5626683
- DOI: 10.1038/s41467-017-00783-2
Molecular architecture of the PBP2-MreC core bacterial cell wall synthesis complex
Abstract
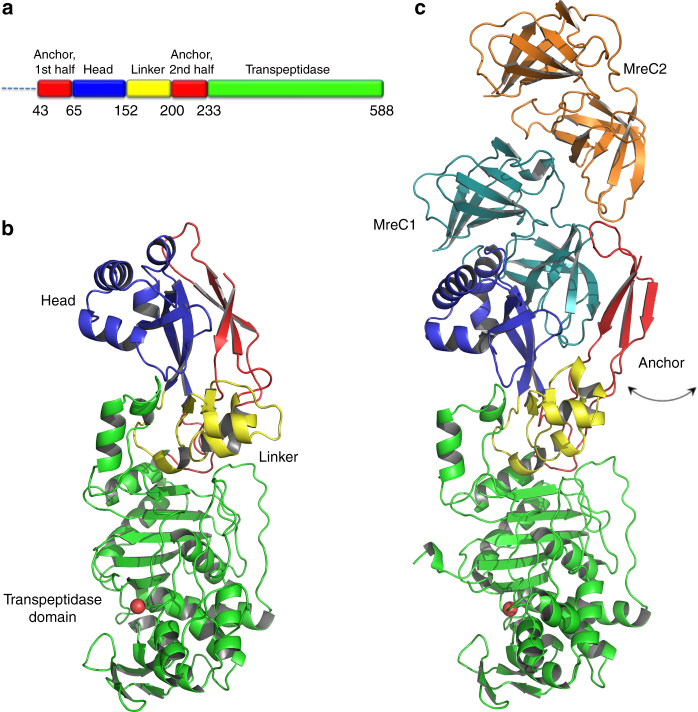
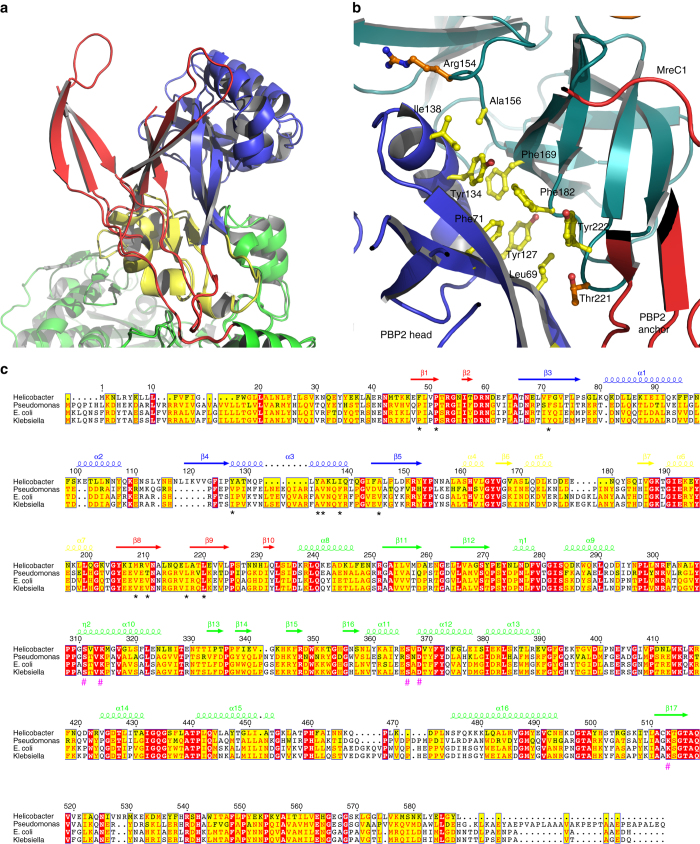
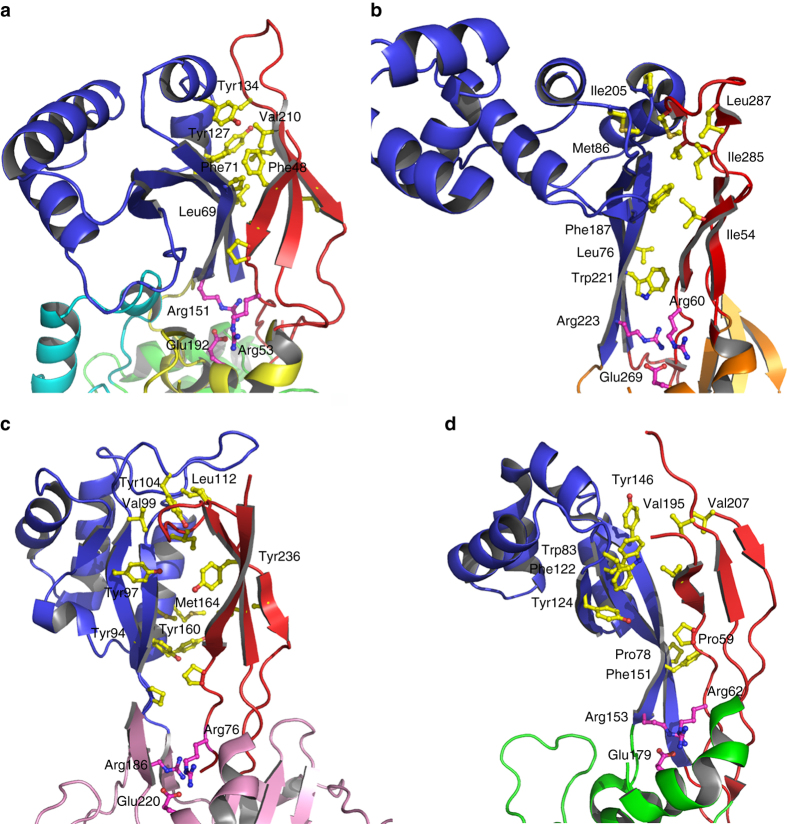
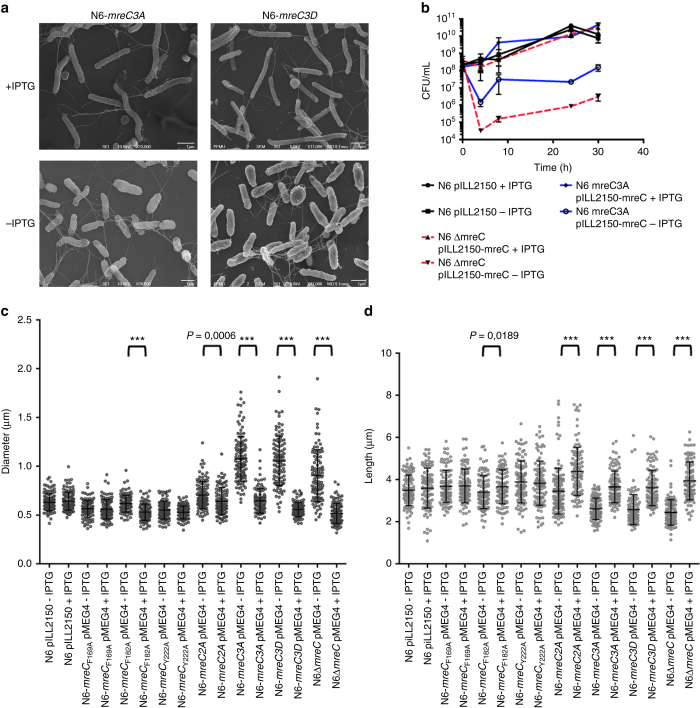
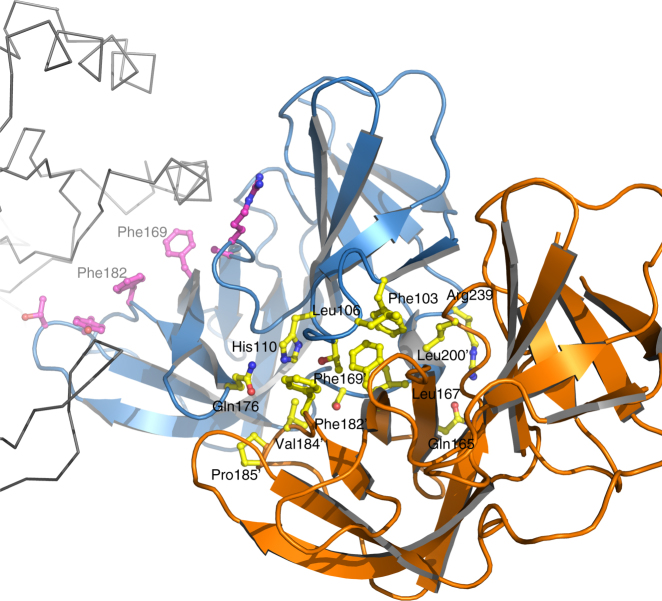
Bacterial cell wall biosynthesis is an essential process that requires the coordinated activity of peptidoglycan biosynthesis enzymes within multi-protein complexes involved in cell division (the "divisome") and lateral wall growth (the "elongasome"). MreC is a structural protein that serves as a platform during wall elongation, scaffolding other essential peptidoglycan biosynthesis macromolecules, such as penicillin-binding proteins. Despite the importance of these multi-partite complexes, details of their architecture have remained elusive due to the transitory nature of their interactions. Here, we present the crystal structures of the soluble PBP2:MreC core elongasome complex from Helicobacter pylori, and of uncomplexed PBP2. PBP2 recognizes the two-winged MreC molecule upon opening of its N-terminal region, revealing a hydrophobic zipper that serves as binding platform. The PBP2:MreC interface is essential both for protein recognition in vitro and maintenance of bacterial shape and growth. This work allows visualization as to how peptidoglycan machinery proteins are scaffolded, revealing interaction regions that could be targeted by tailored inhibitors.Bacterial wall biosynthesis is a complex process that requires the coordination of multiple enzymes. Here, the authors structurally characterize the PBP2:MreC complex involved in peptidoglycan elongation and cross-linking, and demonstrate that its disruption leads to loss of H. pylori shape and inability to sustain growth.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
