

Engineering Camelina sativa (L.) Crantz for enhanced oil and seed yields by combining diacylglycerol acyltransferase1 and glycerol-3-phosphate dehydrogenase expression
- PMID: 28975735
- PMCID: PMC5902773
- DOI: 10.1111/pbi.12847
Engineering Camelina sativa (L.) Crantz for enhanced oil and seed yields by combining diacylglycerol acyltransferase1 and glycerol-3-phosphate dehydrogenase expression
Abstract
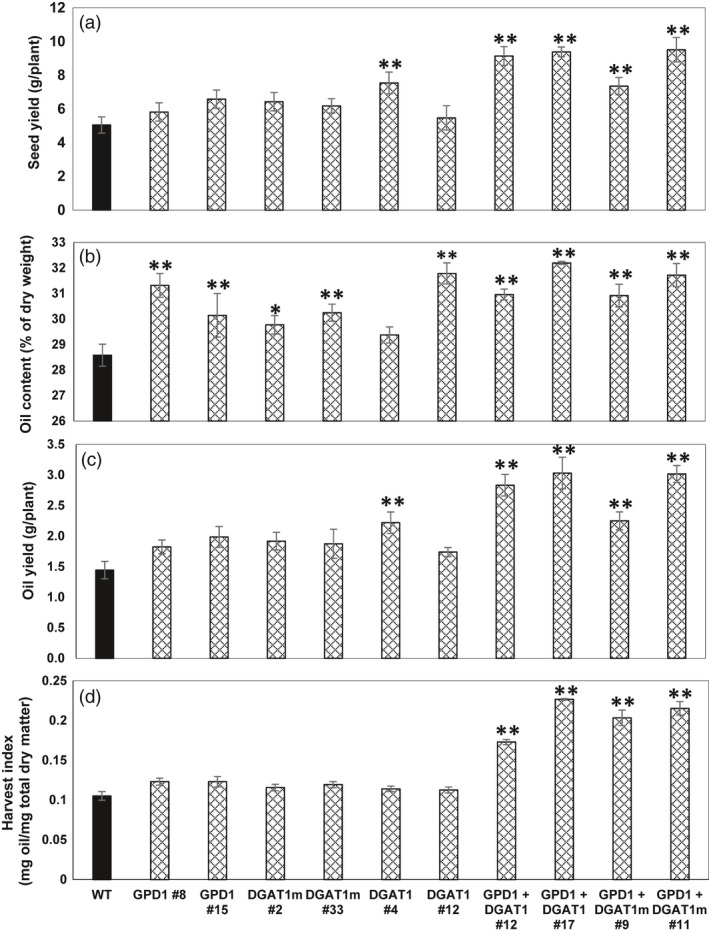
Plant seed oil-based liquid transportation fuels (i.e., biodiesel and green diesel) have tremendous potential as environmentally, economically and technologically feasible alternatives to petroleum-derived fuels. Due to their nutritional and industrial importance, one of the major objectives is to increase the seed yield and oil production of oilseed crops via biotechnological approaches. Camelina sativa, an emerging oilseed crop, has been proposed as an ideal crop for biodiesel and bioproduct applications. Further increase in seed oil yield by increasing the flux of carbon from increased photosynthesis into triacylglycerol (TAG) synthesis will make this crop more profitable. To increase the oil yield, we engineered Camelina by co-expressing the Arabidopsis thaliana (L.) Heynh. diacylglycerol acyltransferase1 (DGAT1) and a yeast cytosolic glycerol-3-phosphate dehydrogenase (GPD1) genes under the control of seed-specific promoters. Plants co-expressing DGAT1 and GPD1 exhibited up to 13% higher seed oil content and up to 52% increase in seed mass compared to wild-type plants. Further, DGAT1- and GDP1-co-expressing lines showed significantly higher seed and oil yields on a dry weight basis than the wild-type controls or plants expressing DGAT1 and GPD1 alone. The oil harvest index (g oil per g total dry matter) for DGTA1- and GPD1-co-expressing lines was almost twofold higher as compared to wild type and the lines expressing DGAT1 and GPD1 alone. Therefore, combining the overexpression of TAG biosynthetic genes, DGAT1 and GPD1, appears to be a positive strategy to achieve a synergistic effect on the flux through the TAG synthesis pathway, and thereby further increase the oil yield.
Keywords: Camelina sativa; biofuels; lipid metabolism; metabolic engineering; triacylglycerols.
© 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Figures






Similar articles
-
Born of frustration: the emergence of Camelina sativa as a platform for lipid biotechnology.Plant Physiol. 2025 Feb 7;197(2):kiaf009. doi: 10.1093/plphys/kiaf009. Plant Physiol. 2025. PMID: 39813144 Free PMC article. Review.
-
Comparative transcriptome and metabolome analysis suggests bottlenecks that limit seed and oil yields in transgenic Camelina sativa expressing diacylglycerol acyltransferase 1 and glycerol-3-phosphate dehydrogenase.Biotechnol Biofuels. 2018 Dec 19;11:335. doi: 10.1186/s13068-018-1326-2. eCollection 2018. Biotechnol Biofuels. 2018. PMID: 30574188 Free PMC article.
-
Camelina sativa: An ideal platform for the metabolic engineering and field production of industrial lipids.Biochimie. 2016 Jan;120:9-16. doi: 10.1016/j.biochi.2015.06.009. Epub 2015 Jun 21. Biochimie. 2016. PMID: 26107412 Review.
-
Increasing seed oil content in oil-seed rape (Brassica napus L.) by over-expression of a yeast glycerol-3-phosphate dehydrogenase under the control of a seed-specific promoter.Plant Biotechnol J. 2007 May;5(3):431-41. doi: 10.1111/j.1467-7652.2007.00252.x. Plant Biotechnol J. 2007. PMID: 17430545
-
Overexpression of the Phosphatidylcholine:DiacylglycerolCholinephosphotransferase (PDCT) gene increases carbon flux toward triacylglycerol (TAG) synthesis in Camelinasativa seeds.Plant Physiol Biochem. 2024 Mar;208:108470. doi: 10.1016/j.plaphy.2024.108470. Epub 2024 Feb 24. Plant Physiol Biochem. 2024. PMID: 38422576
Cited by
-
Increased Cuticle Waxes by Overexpression of WSD1 Improves Osmotic Stress Tolerance in Arabidopsis thaliana and Camelina sativa.Int J Mol Sci. 2021 May 13;22(10):5173. doi: 10.3390/ijms22105173. Int J Mol Sci. 2021. PMID: 34068347 Free PMC article.
-
Seeds as oil factories.Plant Reprod. 2018 Sep;31(3):213-235. doi: 10.1007/s00497-018-0325-6. Epub 2018 Feb 10. Plant Reprod. 2018. PMID: 29429143 Review.
-
Born of frustration: the emergence of Camelina sativa as a platform for lipid biotechnology.Plant Physiol. 2025 Feb 7;197(2):kiaf009. doi: 10.1093/plphys/kiaf009. Plant Physiol. 2025. PMID: 39813144 Free PMC article. Review.
-
Genome-wide profiling of bZIP transcription factors in Camelina sativa: implications for development and stress response.BMC Genom Data. 2024 Oct 14;25(1):88. doi: 10.1186/s12863-024-01270-6. BMC Genom Data. 2024. PMID: 39402491 Free PMC article.
-
Genome-Wide Identification and Expression Analysis of Fatty Acid Desaturase (FAD) Genes in Camelina sativa (L.) Crantz.Int J Mol Sci. 2022 Nov 22;23(23):14550. doi: 10.3390/ijms232314550. Int J Mol Sci. 2022. PMID: 36498878 Free PMC article.
References
-
- An, D. and Suh, M.C. (2015) Overexpression of Arabidopsis WRI1 enhanced seed mass and storage oil content in Camelina sativa. Plant Biotechnol. Rep. 9, 137.
-
- Baud, S. and Lepiniec, L. (2010) Physiological and developmental regulation of seed oil production. Prog. Lipid Res. 49, 235–249. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
