

Involvement of G-triplex and G-hairpin in the multi-pathway folding of human telomeric G-quadruplex
- PMID: 28977514
- PMCID: PMC5737514
- DOI: 10.1093/nar/gkx766
Involvement of G-triplex and G-hairpin in the multi-pathway folding of human telomeric G-quadruplex
Abstract
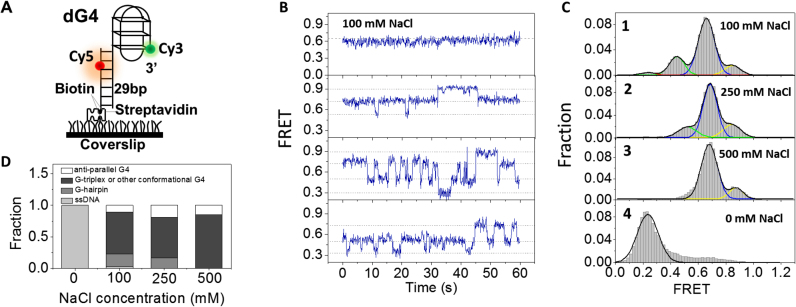
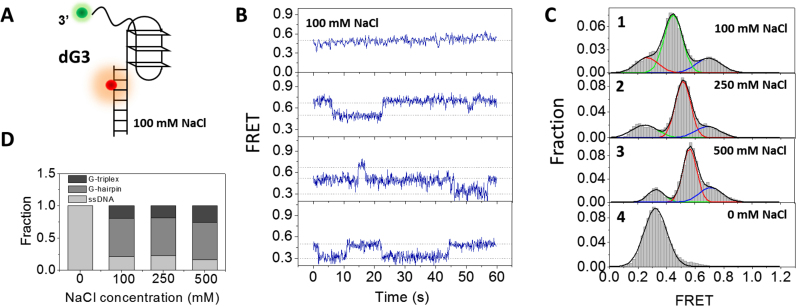
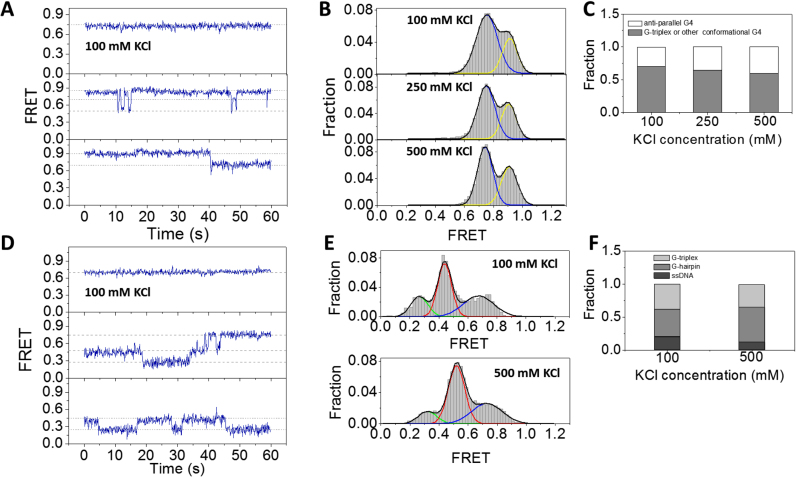
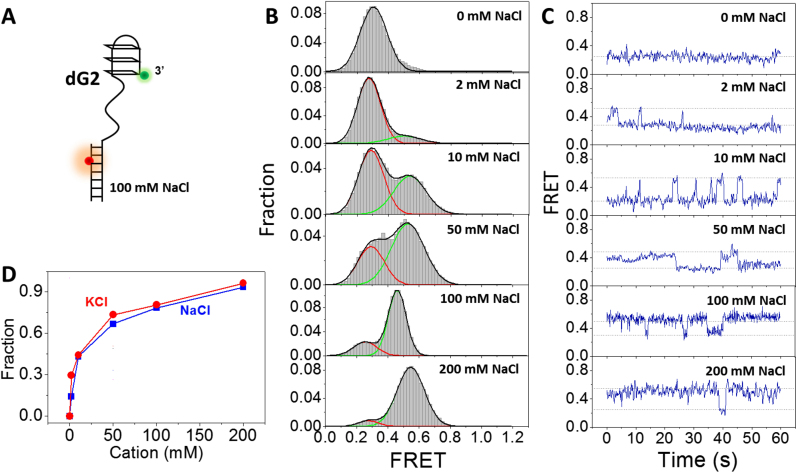
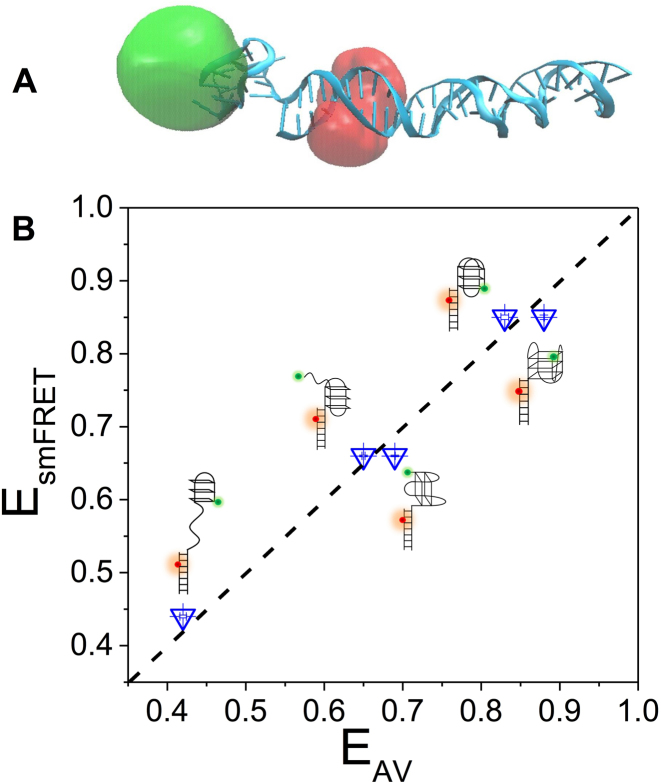
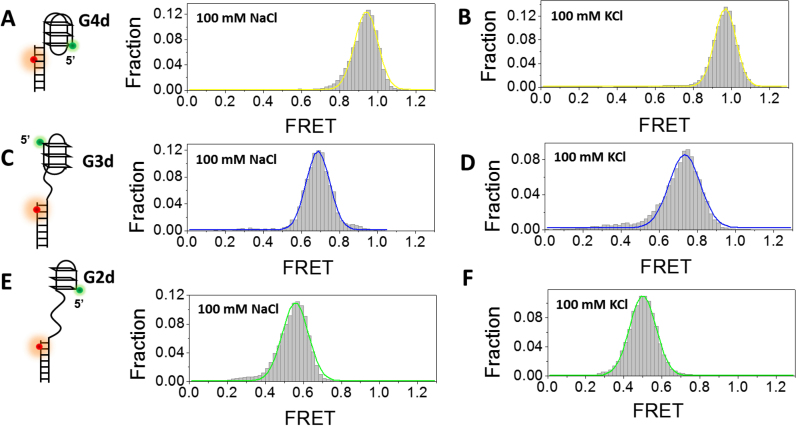
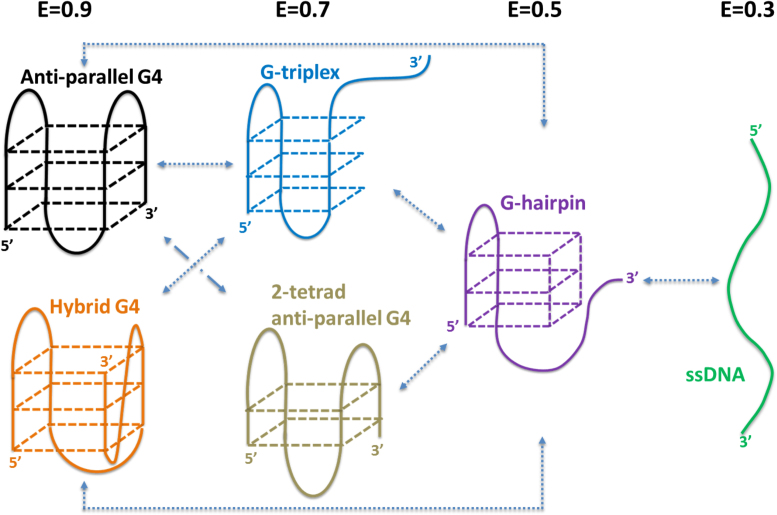
G-quadruplex (G4) can be formed by G-rich DNA sequences that are widely distributed throughout the human genome. Although G-triplex and G-hairpin have been proposed as G4 folding intermediates, their formation still requires further investigation by experiments. Here, we employed single-molecule FRET to characterize the folding dynamics of G4 from human telomeric sequence. First, we observed four states during G4 folding initially assigned to be anti-parallel G4, G-triplex, G-hairpin and unfolded ssDNA. Then we constructed putative intra-strand G-triplex, G-hairpin structures and confirmed their existences in both NaCl and KCl. Further studies revealed those structures are going through dynamic transitions between different states and show relatively weak dependence on cations, unlike G4. Based on those results and molecular dynamics simulations, we proposed a multi-pathway folding mechanism for human telomeric G4. The present work may shed new light on our current understanding about the existence and stability of G4 intermediate states.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
