

Subdomain 2, Not the Transmembrane Domain, Determines the Dimerization Partner of Growth Hormone Receptor and Prolactin Receptor
- PMID: 28977606
- PMCID: PMC5659695
- DOI: 10.1210/en.2017-00469
Subdomain 2, Not the Transmembrane Domain, Determines the Dimerization Partner of Growth Hormone Receptor and Prolactin Receptor
Abstract
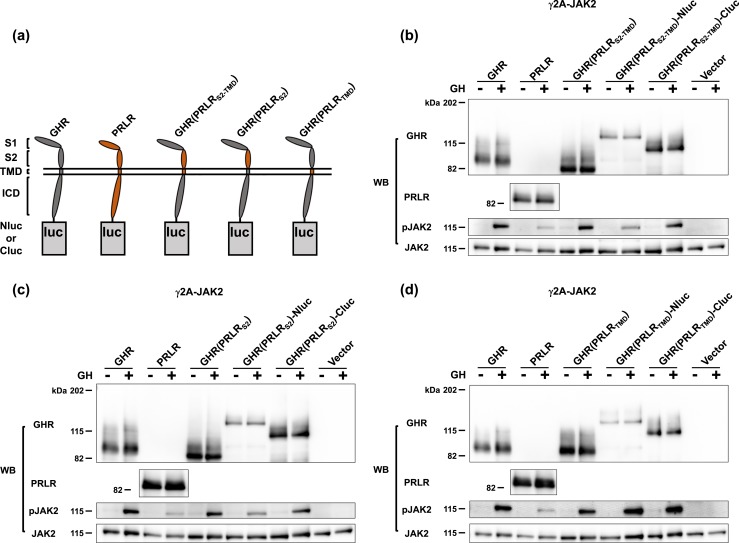
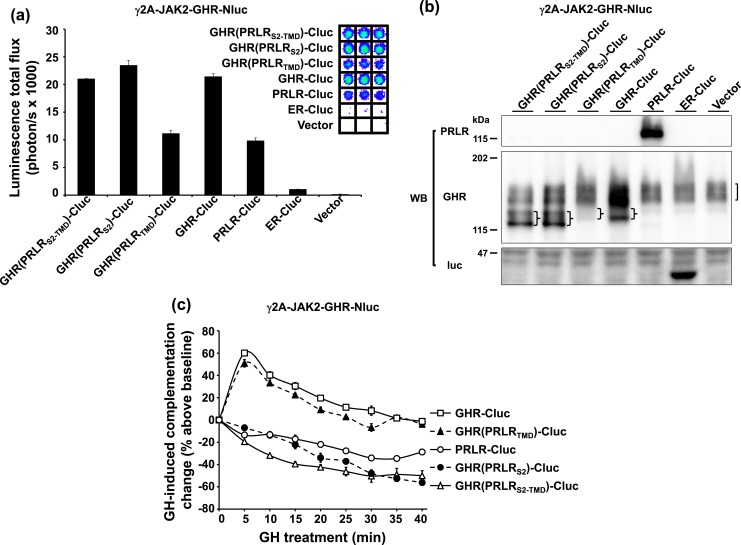
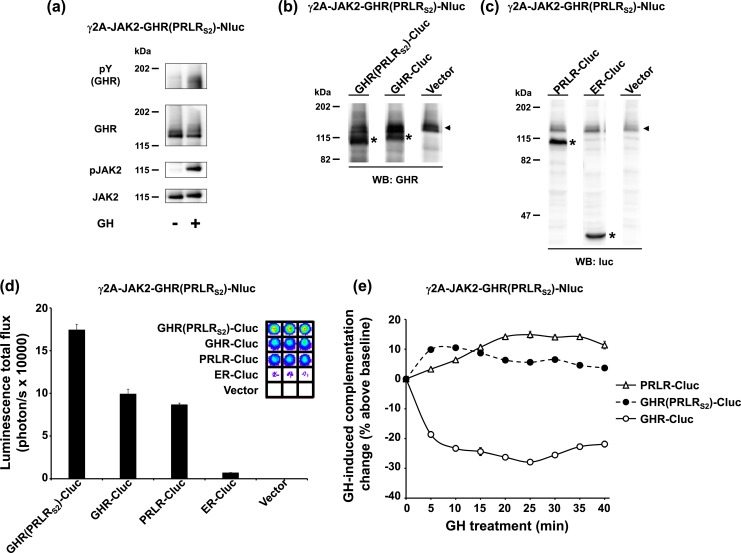
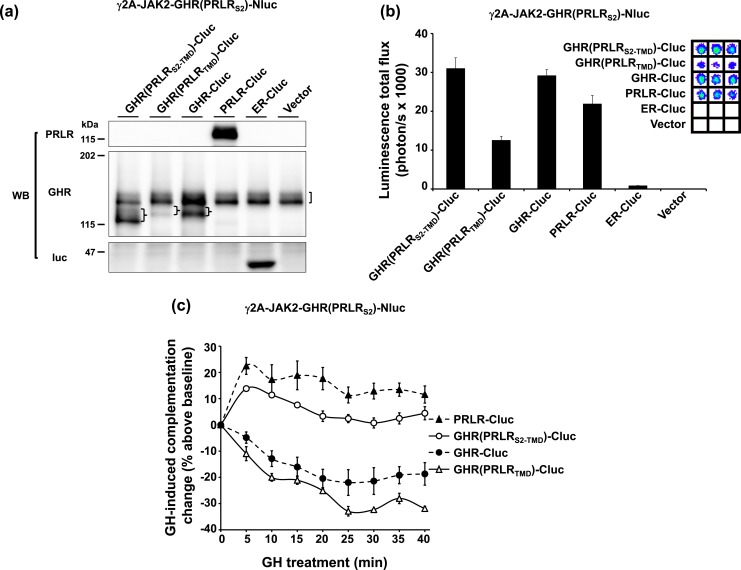
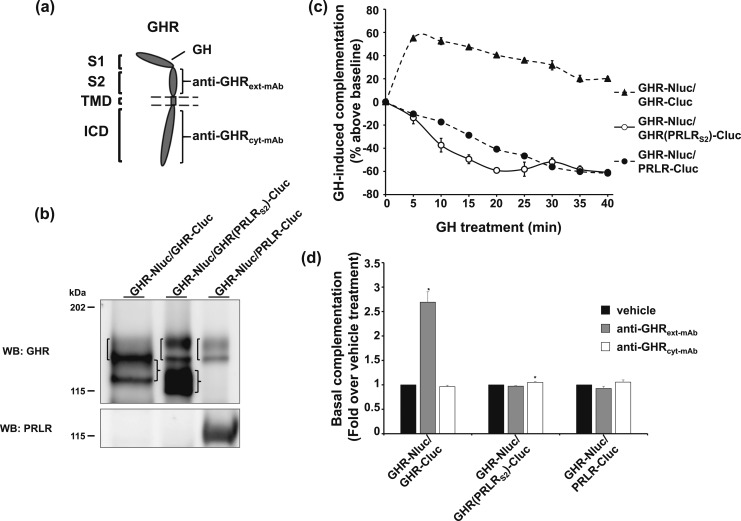
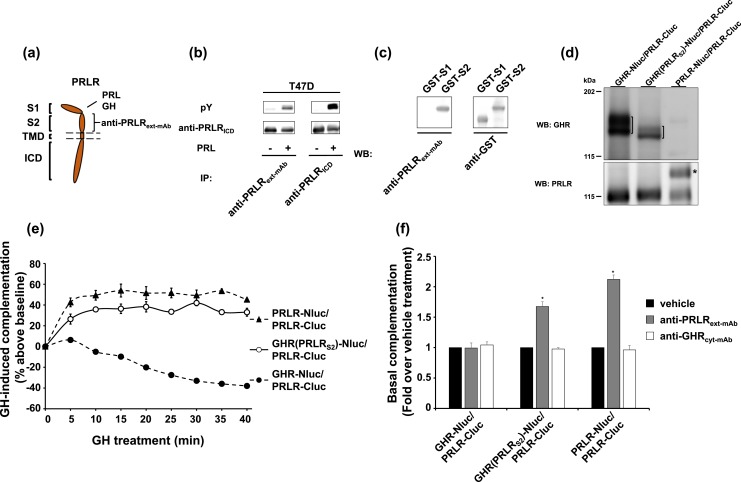
Growth hormone receptor (GHR) and prolactin (PRL) receptor (PRLR) are homologous transmembrane class I cytokine receptors. In humans, GH interacts with GHR homodimers or PRLR homodimers and PRL interacts with only PRLR homodimers to promote signaling. In human breast cancer cells endogenously expressing both receptors, GHR and PRLR specifically coimmunoprecipitate. We previously devised a split luciferase complementation assay to study GHR and PRLR assemblages. In this technique, firefly luciferase is split into two fragments (N- and C-terminal fragments of the luciferase), each without enzyme activity and tethered to the tails of two receptors. The fragments restore luciferase activity when brought close to each other by the receptors. Real-time ligand-induced complementation changes reflect the arrangement of receptors and indicate that GHR/PRLR is arranged as a heteromultimer comprised of GHR-GHR homodimers and PRLR-PRLR homodimers. We now dissect determinants for GHR and PRLR homodimerization versus heteroassociation. GHR and PRLR have extracellular domains comprised of the ligand-binding N-terminal subdomain 1 and a membrane-proximal subdomain 2 (S2), which fosters receptor-receptor contact. Based on previous studies of S2 versus the transmembrane domain (TMD) in GHR dimerization, we constructed GHR(PRLRS2), GHR(PRLRS2-TMD), and GHR(PRLRTMD), replacing GHR's S2 alone, S2 plus TMD, and TMD alone with PRLR's counterpart. We tested by complementation the ability of these chimeras and GHR or PRLR to homodimerize or heteroassociate. Comparing various combinations, we found GHR(PRLRS2) and GHR(PRLRS2-TMD) behaved as PRLR, whereas GHR(PRLRTMD) behaved as GHR regarding their dimerization partners. We conclude that S2 of GHR and PRLR, rather than their TMDs, determines their dimerization partner.
Copyright © 2017 Endocrine Society.
Figures
Similar articles
-
GHR/PRLR Heteromultimer Is Composed of GHR Homodimers and PRLR Homodimers.Mol Endocrinol. 2016 May;30(5):504-17. doi: 10.1210/me.2015-1319. Epub 2016 Mar 22. Mol Endocrinol. 2016. PMID: 27003442 Free PMC article.
-
The role of prolactin receptor in GH signaling in breast cancer cells.Mol Endocrinol. 2013 Feb;27(2):266-79. doi: 10.1210/me.2012-1297. Epub 2012 Nov 28. Mol Endocrinol. 2013. PMID: 23192981 Free PMC article.
-
Growth hormone signaling in human T47D breast cancer cells: potential role for a growth hormone receptor-prolactin receptor complex.Mol Endocrinol. 2011 Apr;25(4):597-610. doi: 10.1210/me.2010-0255. Epub 2011 Feb 10. Mol Endocrinol. 2011. PMID: 21310852 Free PMC article.
-
The role of prolactin and growth hormone in breast cancer.Oncogene. 2000 Feb 21;19(8):1072-6. doi: 10.1038/sj.onc.1203349. Oncogene. 2000. PMID: 10713692 Review.
-
Growth hormone (GH) binding and effects of GH analogs in transgenic mice.Proc Soc Exp Biol Med. 1994 Jul;206(3):190-4. doi: 10.3181/00379727-206-43740. Proc Soc Exp Biol Med. 1994. PMID: 8016152 Review.
Cited by
-
The Relevant Participation of Prolactin in the Genesis and Progression of Gynecological Cancers.Front Endocrinol (Lausanne). 2021 Oct 21;12:747810. doi: 10.3389/fendo.2021.747810. eCollection 2021. Front Endocrinol (Lausanne). 2021. PMID: 34745013 Free PMC article. Review.
-
Growth Hormone Receptor Mutations Related to Individual Dwarfism.Int J Mol Sci. 2018 May 10;19(5):1433. doi: 10.3390/ijms19051433. Int J Mol Sci. 2018. PMID: 29748515 Free PMC article. Review.
-
The Role of the Small Export Apparatus Protein, SctS, in the Activity of the Type III Secretion System.Front Microbiol. 2019 Nov 13;10:2551. doi: 10.3389/fmicb.2019.02551. eCollection 2019. Front Microbiol. 2019. PMID: 31798543 Free PMC article.
-
A Review on Sex Steroid Hormone Estrogen Receptors in Mammals and Fish.Int J Endocrinol. 2020 Feb 7;2020:5386193. doi: 10.1155/2020/5386193. eCollection 2020. Int J Endocrinol. 2020. PMID: 32089683 Free PMC article. Review.
-
Growth hormone (GH) receptor (GHR)-specific inhibition of GH-Induced signaling by soluble IGF-1 receptor (sol IGF-1R).Mol Cell Endocrinol. 2019 Jul 15;492:110445. doi: 10.1016/j.mce.2019.05.004. Epub 2019 May 14. Mol Cell Endocrinol. 2019. PMID: 31100495 Free PMC article.
References
-
- Yang N, Wang X, Jiang J, Frank SJ. Role of the growth hormone (GH) receptor transmembrane domain in receptor predimerization and GH-induced activation. Mol Endocrinol. 2007;21(7):1642–1655. - PubMed
-
- Brown RJ, Adams JJ, Pelekanos RA, Wan Y, McKinstry WJ, Palethorpe K, Seeber RM, Monks TA, Eidne KA, Parker MW, Waters MJ. Model for growth hormone receptor activation based on subunit rotation within a receptor dimer. Nat Struct Mol Biol. 2005;12(9):814–821. - PubMed
-
- Biener-Ramanujan E, Ramanujan VK, Herman B, Gertler A. Spatio-temporal kinetics of growth hormone receptor signaling in single cells using FRET microscopy. Growth Horm IGF Res. 2006;16(4):247–257. - PubMed
-
- Brooks AJ, Dai W, O’Mara ML, Abankwa D, Chhabra Y, Pelekanos RA, Gardon O, Tunny KA, Blucher KM, Morton CJ, Parker MW, Sierecki E, Gambin Y, Gomez GA, Alexandrov K, Wilson IA, Doxastakis M, Mark AE, Waters MJ. Mechanism of activation of protein kinase JAK2 by the growth hormone receptor. Science. 2014;344(6185):1249783. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
