

PP32 and SET/TAF-Iβ proteins regulate the acetylation of newly synthesized histone H4
- PMID: 28977641
- PMCID: PMC5714232
- DOI: 10.1093/nar/gkx775
PP32 and SET/TAF-Iβ proteins regulate the acetylation of newly synthesized histone H4
Abstract
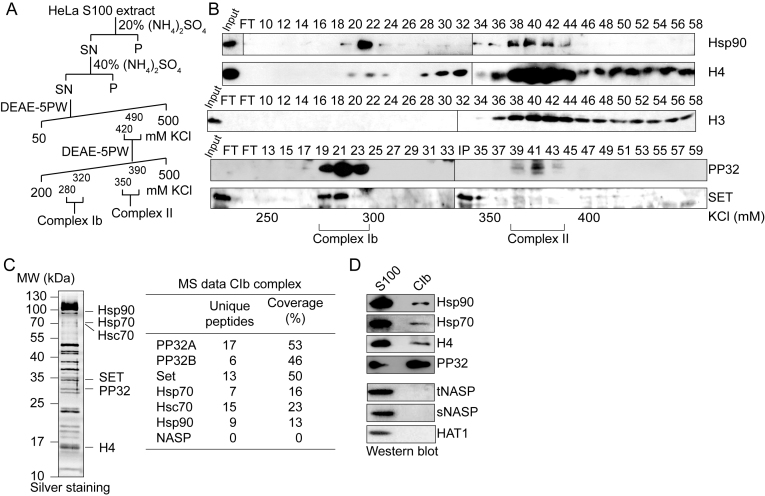
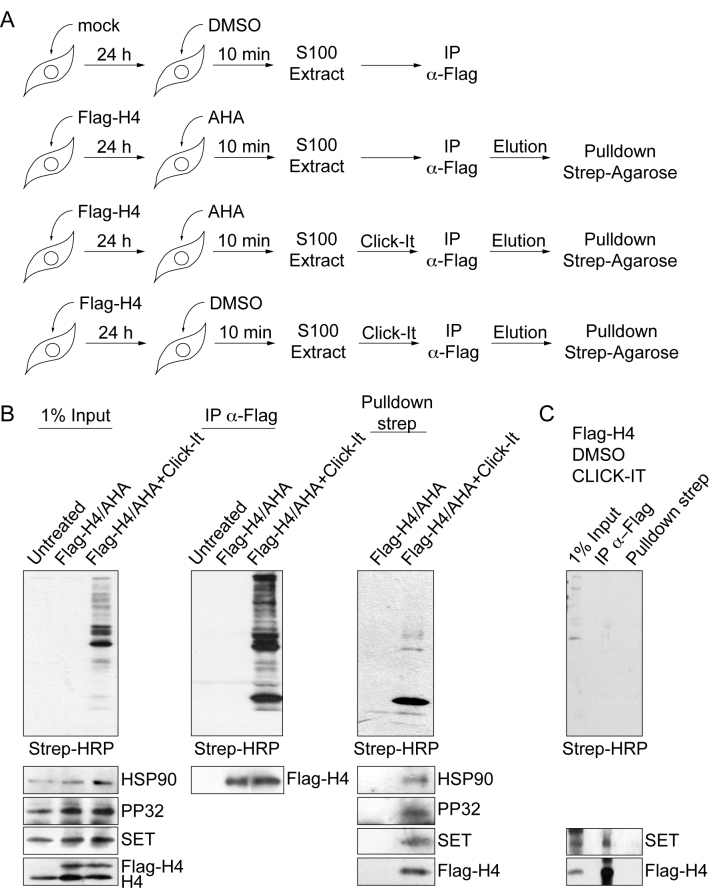
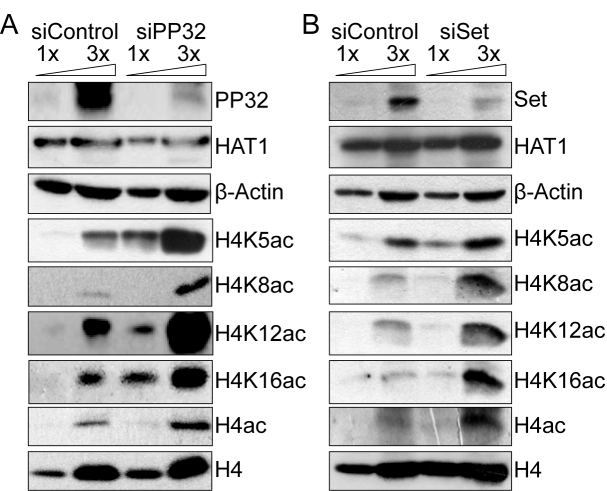
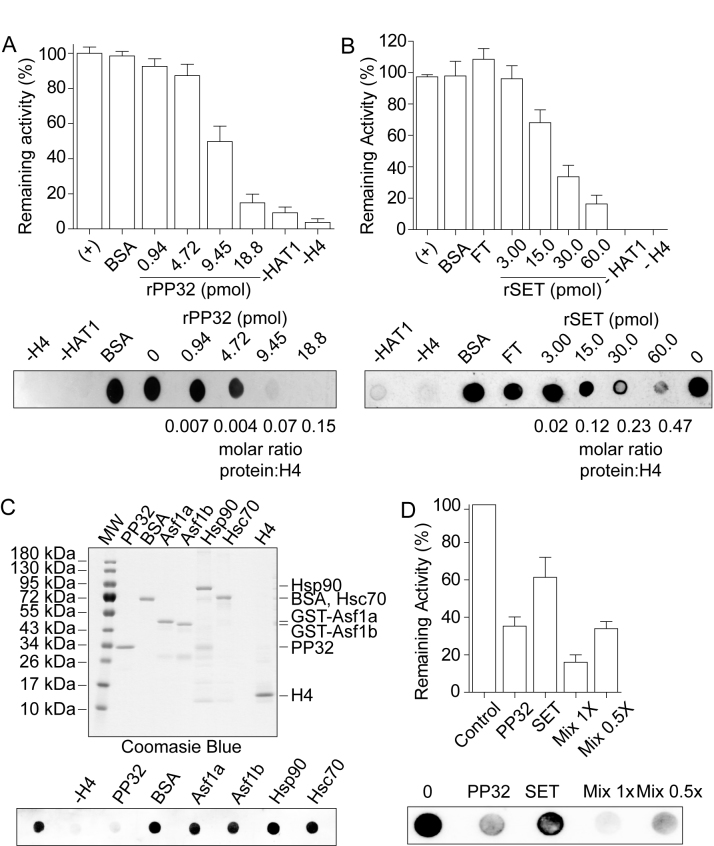
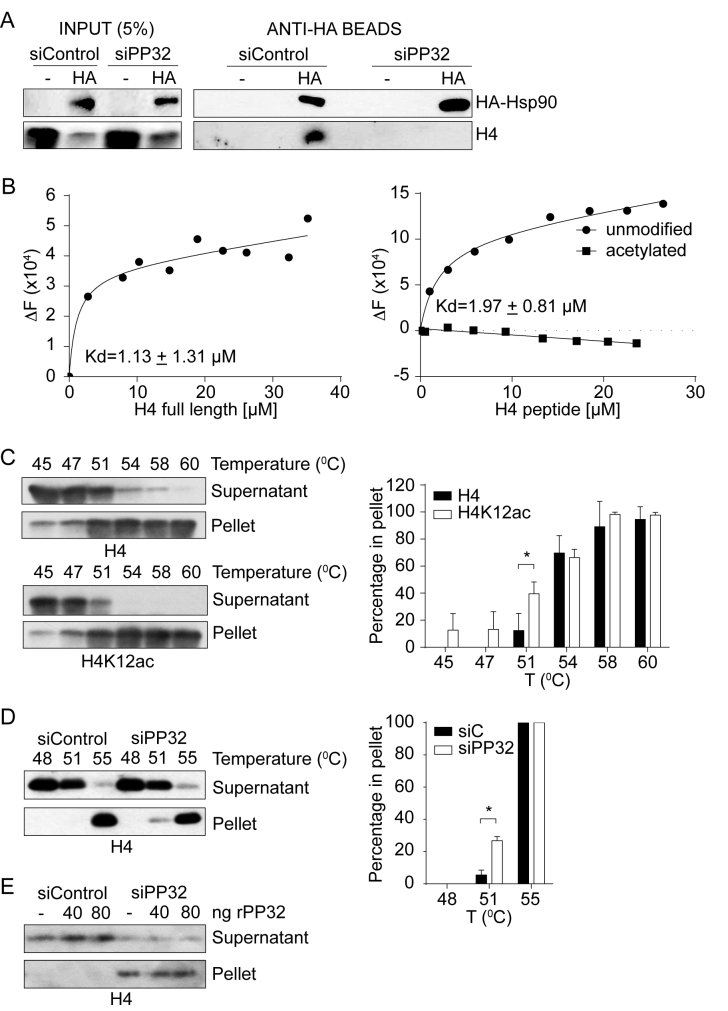
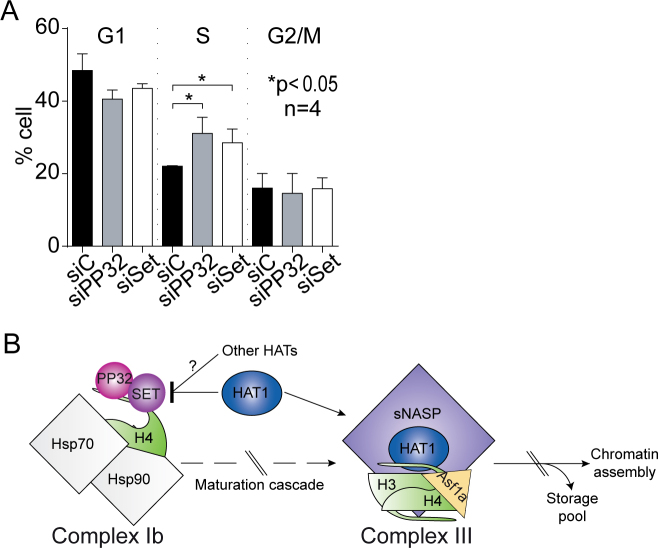
Newly synthesized histones H3 and H4 undergo a cascade of maturation steps to achieve proper folding and to establish post-translational modifications prior to chromatin deposition. Acetylation of H4 on lysines 5 and 12 by the HAT1 acetyltransferase is observed late in the histone maturation cascade. A key question is to understand how to establish and regulate the distinct timing of sequential modifications and their biological significance. Here, we perform proteomic analysis of the newly synthesized histone H4 complex at the earliest time point in the cascade. In addition to known binding partners Hsp90 and Hsp70, we also identify for the first time two subunits of the histone acetyltransferase inhibitor complex (INHAT): PP32 and SET/TAF-Iβ. We show that both proteins function to prevent HAT1-mediated H4 acetylation in vitro. When PP32 and SET/TAF-Iβ protein levels are down-regulated in vivo, we detect hyperacetylation on lysines 5 and 12 and other H4 lysine residues. Notably, aberrantly acetylated H4 is less stable and this reduces the interaction with Hsp90. As a consequence, PP32 and SET/TAF-Iβ depleted cells show an S-phase arrest. Our data demonstrate a novel function of PP32 and SET/TAF-Iβ and provide new insight into the mechanisms regulating acetylation of newly synthesized histone H4.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Marzluff W.F., Duronio R.J.. Histone mRNA expression: multiple levels of cell cycle regulation and important developmental consequences. Curr. Opin. Cell Biol. 2002; 14:692–699. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
