

Plant organellar DNA polymerases are replicative and translesion DNA synthesis polymerases
- PMID: 28977655
- PMCID: PMC5737093
- DOI: 10.1093/nar/gkx744
Plant organellar DNA polymerases are replicative and translesion DNA synthesis polymerases
Abstract
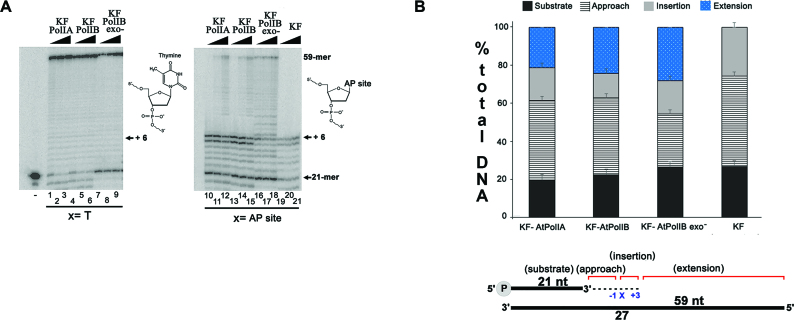
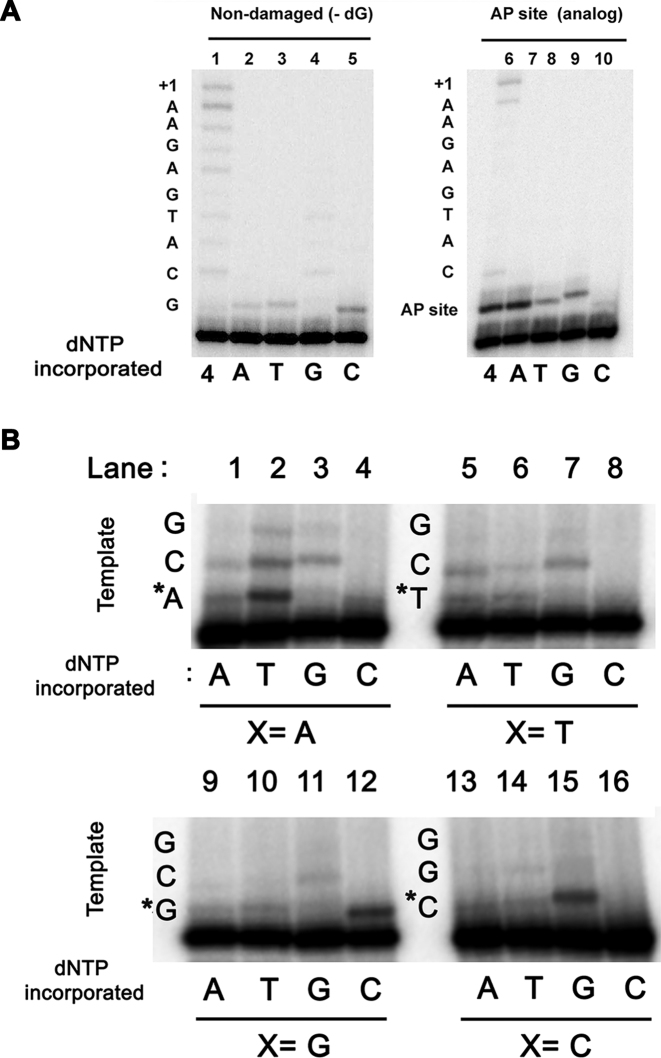
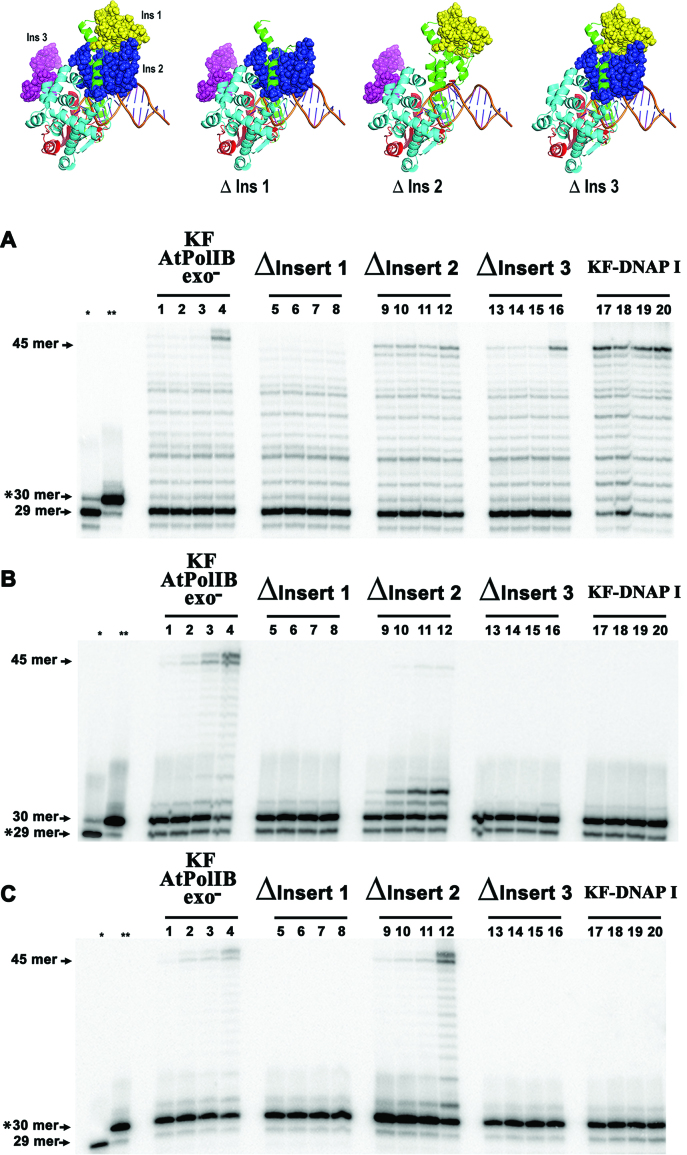
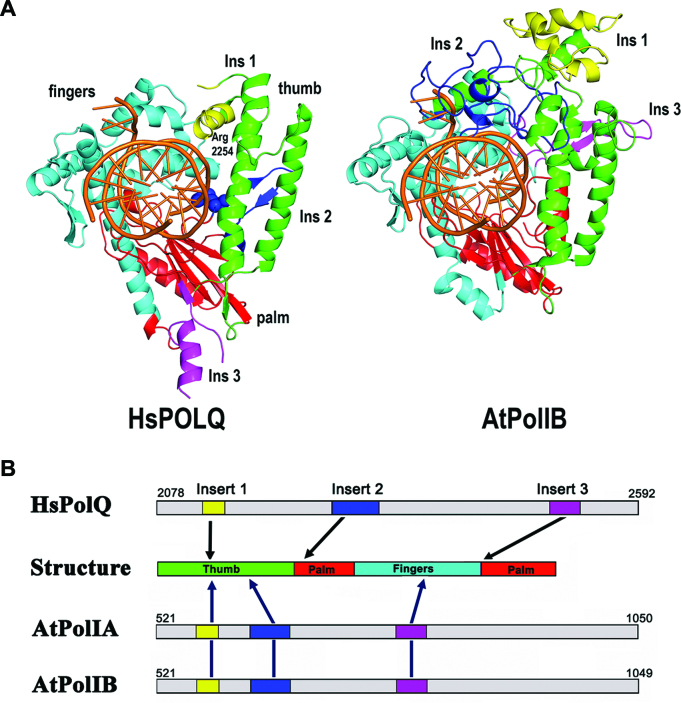
Genomes acquire lesions that can block the replication fork and some lesions must be bypassed to allow survival. The nuclear genome of flowering plants encodes two family-A DNA polymerases (DNAPs), the result of a duplication event, that are the sole DNAPs in plant organelles. These DNAPs, dubbed Plant Organellar Polymerases (POPs), resemble the Klenow fragment of bacterial DNAP I and are not related to metazoan and fungal mitochondrial DNAPs. Herein we report that replicative POPs from the plant model Arabidopsis thaliana (AtPolI) efficiently bypass one the most insidious DNA lesions, an apurinic/apyrimidinic (AP) site. AtPolIs accomplish lesion bypass with high catalytic efficiency during nucleotide insertion and extension. Lesion bypass depends on two unique polymerization domain insertions evolutionarily unrelated to the insertions responsible for lesion bypass by DNAP θ, an analogous lesion bypass polymerase. AtPolIs exhibit an insertion fidelity that ranks between the fidelity of replicative and lesion bypass DNAPs, moderate 3'-5' exonuclease activity and strong strand-displacement. AtPolIs are the first known example of a family-A DNAP evolved to function in both DNA replication and lesion bypass. The lesion bypass capabilities of POPs may be required to prevent replication fork collapse in plant organelles.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Plant organellar DNA polymerases paralogs exhibit dissimilar nucleotide incorporation fidelity.FEBS J. 2018 Nov;285(21):4005-4018. doi: 10.1111/febs.14645. Epub 2018 Sep 11. FEBS J. 2018. PMID: 30152200
-
Identification of a unique insertion in plant organellar DNA polymerases responsible for 5'-dRP lyase and strand-displacement activities: Implications for Base Excision Repair.DNA Repair (Amst). 2018 May;65:1-10. doi: 10.1016/j.dnarep.2018.02.010. Epub 2018 Feb 27. DNA Repair (Amst). 2018. PMID: 29522990
-
Plant organellar DNA polymerases bypass thymine glycol using two conserved lysine residues.Biochem J. 2020 Mar 13;477(5):1049-1059. doi: 10.1042/BCJ20200043. Biochem J. 2020. PMID: 32108856
-
Plant Organellar DNA Polymerases Evolved Multifunctionality through the Acquisition of Novel Amino Acid Insertions.Genes (Basel). 2020 Nov 19;11(11):1370. doi: 10.3390/genes11111370. Genes (Basel). 2020. PMID: 33228188 Free PMC article. Review.
-
Eukaryotic translesion synthesis DNA polymerases: specificity of structure and function.Annu Rev Biochem. 2005;74:317-53. doi: 10.1146/annurev.biochem.74.082803.133250. Annu Rev Biochem. 2005. PMID: 15952890 Review.
Cited by
-
R-loop control and mitochondrial genome stability require the 5'-3' exonuclease/flap endonuclease OEX1.Plant Cell. 2025 May 9;37(5):koaf104. doi: 10.1093/plcell/koaf104. Plant Cell. 2025. PMID: 40324391
-
Plant Organellar MSH1 Is a Displacement Loop-Specific Endonuclease.Plant Cell Physiol. 2024 May 14;65(4):560-575. doi: 10.1093/pcp/pcad112. Plant Cell Physiol. 2024. PMID: 37756637 Free PMC article.
-
Arabidopsis thaliana organelles mimic the T7 phage DNA replisome with specific interactions between Twinkle protein and DNA polymerases Pol1A and Pol1B.BMC Plant Biol. 2019 Jun 6;19(1):241. doi: 10.1186/s12870-019-1854-3. BMC Plant Biol. 2019. PMID: 31170927 Free PMC article.
-
Arabidopsis thaliana PrimPol is a primase and lesion bypass DNA polymerase with the biochemical characteristics to cope with DNA damage in the nucleus, mitochondria, and chloroplast.Sci Rep. 2021 Oct 18;11(1):20582. doi: 10.1038/s41598-021-00151-7. Sci Rep. 2021. PMID: 34663822 Free PMC article.
-
DNA Polymerases for Whole Genome Amplification: Considerations and Future Directions.Int J Mol Sci. 2023 May 26;24(11):9331. doi: 10.3390/ijms24119331. Int J Mol Sci. 2023. PMID: 37298280 Free PMC article. Review.
References
-
- Yokoi M., Ito M., Izumi M., Miyazawa H., Nakai H., Hanaoka F.. Molecular cloning of the cDNA for the catalytic subunit of plant DNA polymerase alpha and its cell-cycle dependent expression. Genes Cells. 1997; 2:695–709. - PubMed
-
- Anderson H.J., Vonarx E.J., Pastushok L., Nakagawa M., Katafuchi A., Gruz P., Di Rubbo A., Grice D.M., Osmond M.J., Sakamoto A.N. et al. . Arabidopsis thaliana Y-family DNA polymerase eta catalyses translesion synthesis and interacts functionally with PCNA2. Plant J. 2008; 55:895–908. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
