

Hyperthermia and associated changes in membrane fluidity potentiate P2X7 activation to promote tumor cell death
- PMID: 28978031
- PMCID: PMC5620171
- DOI: 10.18632/oncotarget.18595
Hyperthermia and associated changes in membrane fluidity potentiate P2X7 activation to promote tumor cell death
Abstract
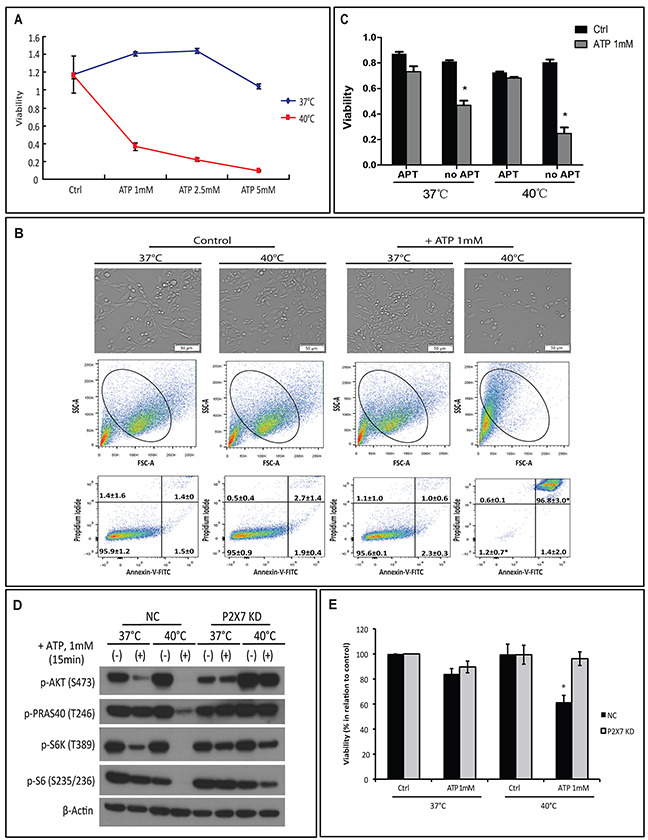
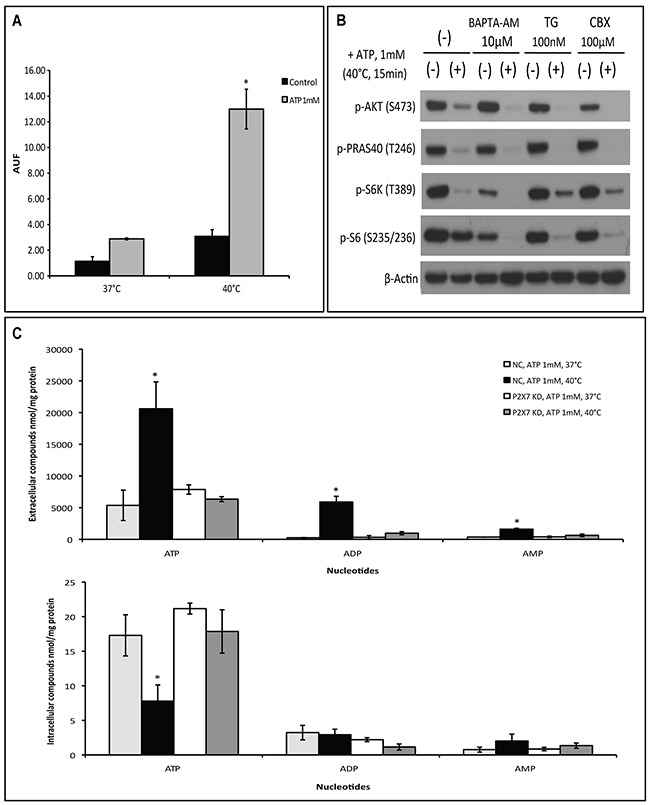
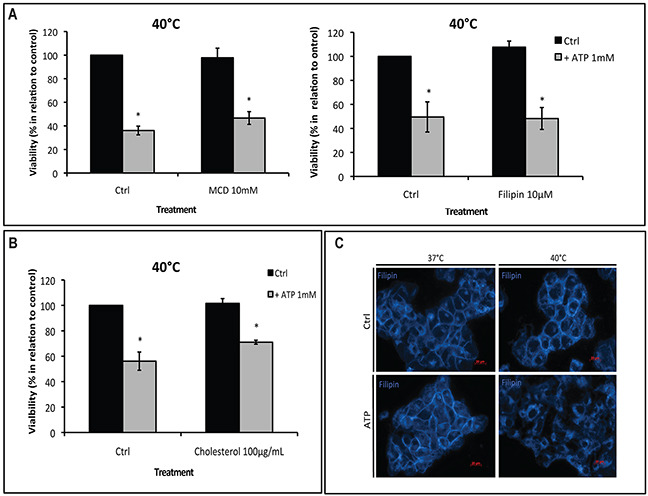
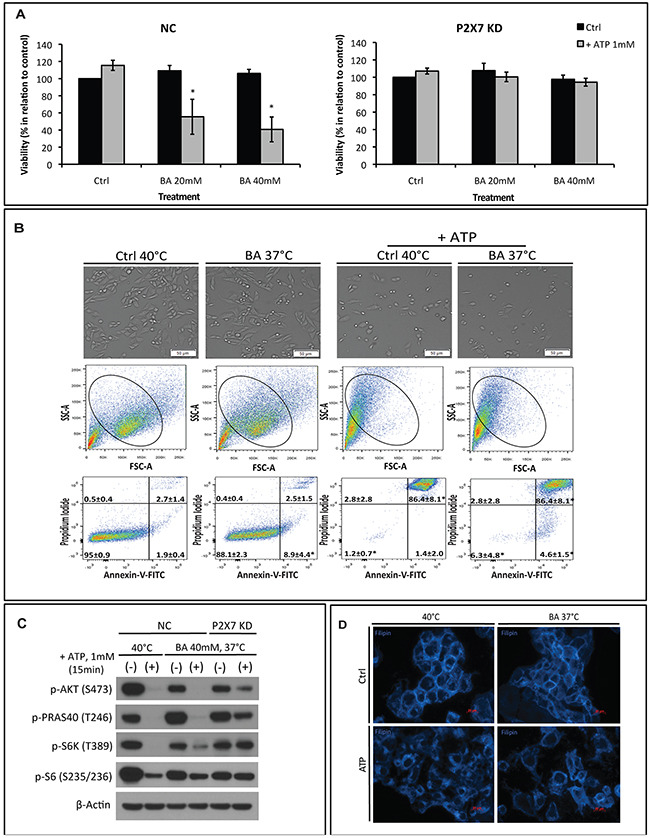
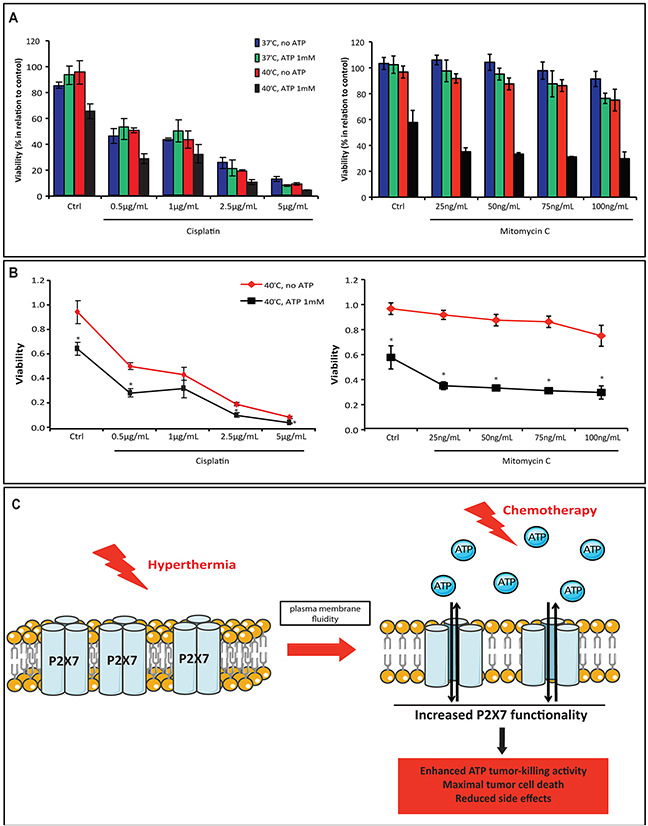
Extracellular ATP (eATP) accumulation within the tumor microenvironment (TME) has the potential to activate purinergic signaling. The eATP evoked signaling effects bolster antitumor immune responses while exerting direct cytotoxicity on tumor cells and vascular endothelial cells, mediated at least in part through P2X7 receptors. Approaches to augment purinergic signaling in TME e.g. by ectonucleotidase CD39 blockade, and/or boosting P2X7 functional responses, might be used as immunomodulatory therapies in cancer treatment. In this study, we delineated the translatable strategy of hyperthermia to demonstrate impacts on P2X7 responsiveness to eATP. Hyperthermia (40°C) was noted to enhance eATP-mediated cytotoxicity on MCA38 colon cancer cells. Increased membrane fluidity induced by hyperthermia boosted P2X7 functionality, potentiating pore opening and modulating downstream AKT/PRAS40/mTOR signaling events. When combined with cisplatin or mitomycin C, hyperthermia and eATP together markedly potentiate cancer cell death. Our data indicate that clinically tolerable hyperthermia with modulated P2X7-purinergic signaling will boost efficacy of conventional cancer treatments.
Keywords: cancer therapy; colon cancer; hyperthermia; membrane fluidity; purinergic signaling.
Conflict of interest statement
CONFLICTS OF INTEREST The authors declare no competing or financial interests.
Figures
References
-
- Roger S, Jelassi B, Couillin I, Pelegrin P, Besson P, Jiang LH. Understanding the roles of the P2X7 receptor in solid tumour progression and therapeutic perspectives. Biochim Biophys Acta. 2015;1848:2584–602. - PubMed
-
- Feng L, Tapper EB, Sun X, Gehring M, Robson SC, Wu Y. In: Frontiers in Anti-Cancer Drug Discovery. Atta-ur-Rahman, Iqbal Choudhary M, editors. Bentham Science Publishers; Beijing, China: 2014. Purinegic Modulation and CD39/ENTPD1 in Cancer; pp. 229–292.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
