

Molecular plasticity and functional enhancements of leg muscles in response to hypergravity in the fruit fly Drosophila melanogaster
- PMID: 28978639
- PMCID: PMC5665435
- DOI: 10.1242/jeb.160523
Molecular plasticity and functional enhancements of leg muscles in response to hypergravity in the fruit fly Drosophila melanogaster
Abstract
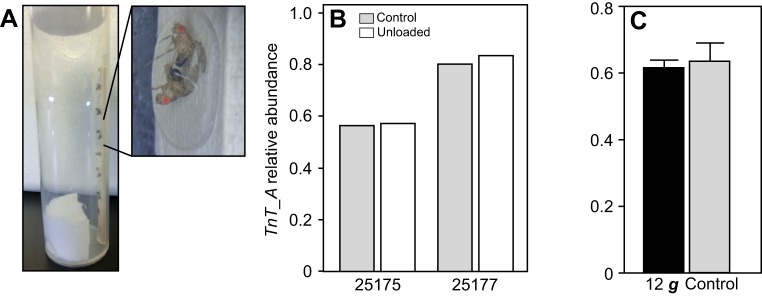
Studies of organismal and tissue biomechanics have clearly demonstrated that musculoskeletal design is strongly dependent on experienced loads, which can vary in the short term, as a result of growth during life history and during the evolution of animal body size. However, how animals actually perceive and make adjustments to their load-bearing musculoskeletal elements that accommodate variation in their body weight is poorly understood. We developed an experimental model system that can be used to start addressing these open questions, and uses hypergravity centrifugation to experimentally manipulate the loads experienced by Drosophila melanogaster We examined effects of this manipulation on leg muscle alternative splicing of the sarcomere gene troponin T (Dmel\up; Fbgn0004169, herein referred to by its synonym TnT), a process that was previously demonstrated to precisely correlate with quantitative variation in body weight in Lepidoptera and rat. In a similar fashion, hypergravity centrifugation caused fast (i.e. within 24 h) changes to fly leg muscle TnT alternative splicing that correlated with body weight variation across eight D. melanogaster lines. Hypergravity treatment also appeared to enhance leg muscle function, as centrifuged flies showed an increased negative geotaxis response and jump ability. Although the identity and location of the sensors and effectors involved remains unknown, our results provide further support for the existence of an evolutionarily conserved mechanism that translates signals that encode body weight into appropriate skeletal muscle molecular and functional responses.
Keywords: Alternative splicing; Body mass; Body weight; Centrifugation; Muscle performance; Musculoskeletal; Troponin T; Weight sensing.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures






Similar articles
-
Body weight-dependent troponin T alternative splicing is evolutionarily conserved from insects to mammals and is partially impaired in skeletal muscle of obese rats.J Exp Biol. 2011 May 1;214(Pt 9):1523-32. doi: 10.1242/jeb.051763. J Exp Biol. 2011. PMID: 21490260 Free PMC article.
-
Cell-autonomous regulation of fast troponin T pre-mRNA alternative splicing in response to mechanical stretch.Am J Physiol Cell Physiol. 2012 Aug 1;303(3):C298-307. doi: 10.1152/ajpcell.00400.2011. Epub 2012 May 16. Am J Physiol Cell Physiol. 2012. PMID: 22592404 Free PMC article.
-
Weight and nutrition affect pre-mRNA splicing of a muscle gene associated with performance, energetics and life history.J Exp Biol. 2008 Dec;211(Pt 23):3653-60. doi: 10.1242/jeb.023903. J Exp Biol. 2008. PMID: 19011203
-
TNNT1, TNNT2, and TNNT3: Isoform genes, regulation, and structure-function relationships.Gene. 2016 May 10;582(1):1-13. doi: 10.1016/j.gene.2016.01.006. Epub 2016 Jan 13. Gene. 2016. PMID: 26774798 Free PMC article. Review.
-
Effects of microgravity and hypergravity on aging and longevity of insects.Korean J Biol Sci. 2000 Sep;4(3):231-7. doi: 10.1080/12265071.2000.9647549. Korean J Biol Sci. 2000. PMID: 12760374 Review.
Cited by
-
Loss of Fic causes progressive neurodegeneration in a Drosophila model of hereditary spastic paraplegia.Biochim Biophys Acta Mol Basis Dis. 2024 Oct;1870(7):167348. doi: 10.1016/j.bbadis.2024.167348. Epub 2024 Jul 8. Biochim Biophys Acta Mol Basis Dis. 2024. PMID: 38986817
-
Automated insect detection and biomass monitoring via AI and electrical field sensor technology.Sci Rep. 2025 Aug 14;15(1):29858. doi: 10.1038/s41598-025-15613-5. Sci Rep. 2025. PMID: 40813816 Free PMC article.
-
The effects of genetic background on exercise performance in Drosophila.Fly (Austin). 2020 Mar-Dec;14(1-4):80-92. doi: 10.1080/19336934.2020.1835329. Epub 2020 Oct 26. Fly (Austin). 2020. PMID: 33100141 Free PMC article.
References
-
- Alexander R. M. N. (1976). Estimates of speeds of dinosaurs. Nature 261, 129-130. 10.1038/261129a0 - DOI
-
- Alexander R. M. N. (1985). The maximum forces exerted by animals. J. Exp. Biol. 115, 231-238. - PubMed
-
- Alexander R. M. N. (2003). Principles of Animal Locomotion. Princeton, NJ: Princeton University Press.
-
- Alexander R. M. N. and Jayes A. S. (1983). A dynamic similarity hypothesis for the gaits of quadrupedal mammals. J. Zool. 201, 135-152. 10.1111/j.1469-7998.1983.tb04266.x - DOI
-
- Atherton P. J., Szewczyk N. J., Selby A., Rankin D., Hillier K., Smith K., Rennie M. J. and Loughna P. T. (2009). Cyclic stretch reduces myofibrillar protein synthesis despite increases in FAK and anabolic signalling in L6 cells. J. Physiol. 587, 3719-3727. 10.1113/jphysiol.2009.169854 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
