

Processing of Potato Spindle Tuber Viroid RNAs in Yeast, a Nonconventional Host
- PMID: 28978701
- PMCID: PMC5709609
- DOI: 10.1128/JVI.01078-17
Processing of Potato Spindle Tuber Viroid RNAs in Yeast, a Nonconventional Host
Abstract
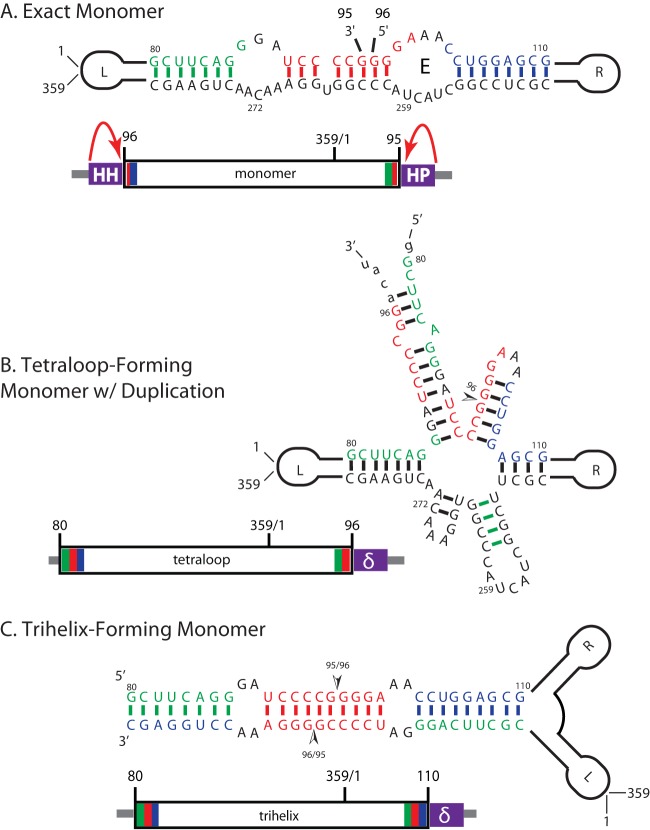
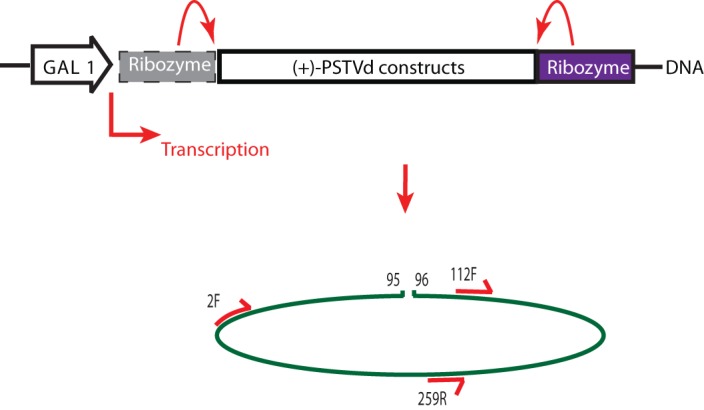
Potato spindle tuber viroid (PSTVd) is a circular, single-stranded, noncoding RNA plant pathogen that is a useful model for studying the processing of noncoding RNA in eukaryotes. Infective PSTVd circles are replicated via an asymmetric rolling circle mechanism to form linear multimeric RNAs. An endonuclease cleaves these into monomers, and a ligase seals these into mature circles. All eukaryotes may have such enzymes for processing noncoding RNA. As a test, we investigated the processing of three PSTVd RNA constructs in the yeast Saccharomyces cerevisiae Of these, only one form, a construct that adopts a previously described tetraloop-containing conformation (TL), produces circles. TL has 16 nucleotides of the 3' end duplicated at the 5' end and a 3' end produced by self-cleavage of a delta ribozyme. The other two constructs, an exact monomer flanked by ribozymes and a trihelix-forming RNA with requisite 5' and 3' duplications, do not produce circles. The TL circles contain nonnative nucleotides resulting from the 3' end created by the ribozyme and the 5' end created from an endolytic cleavage by yeast at a site distinct from where potato enzymes cut these RNAs. RNAs from all three transcripts are cleaved in places not on path for circle formation, likely representing RNA decay. We propose that these constructs fold into distinct RNA structures that interact differently with host cell RNA metabolism enzymes, resulting in various susceptibilities to degradation versus processing. We conclude that PSTVd RNA is opportunistic and may use different processing pathways in different hosts.IMPORTANCE In higher eukaryotes, the majority of transcribed RNAs do not encode proteins. These noncoding RNAs are responsible for mRNA regulation, control of the expression of regulatory microRNAs, sensing of changes in the environment by use of riboswitches (RNAs that change shape in response to environmental signals), catalysis, and more roles that are still being uncovered. Some of these functions may be remnants from the RNA world and, as such, would be part of the evolutionary past of all forms of modern life. Viroids are noncoding RNAs that can cause disease in plants. Since they encode no proteins, they depend on their own RNA and on host proteins for replication and pathogenicity. It is likely that viroids hijack critical host RNA pathways for processing the host's own noncoding RNA. These pathways are still unknown. Elucidating these pathways should reveal new biological functions of noncoding RNA.
Keywords: RNA conformation; RNA processing; host functions; mRNA degradation; noncoding RNA; viroids.
Copyright © 2017 American Society for Microbiology.
Figures

Similar articles
-
Potato Spindle Tuber Viroid Modulates Its Replication through a Direct Interaction with a Splicing Regulator.J Virol. 2018 Sep 26;92(20):e01004-18. doi: 10.1128/JVI.01004-18. Print 2018 Oct 15. J Virol. 2018. PMID: 30068655 Free PMC article.
-
Comprehensive Transcriptome Analyses Reveal that Potato Spindle Tuber Viroid Triggers Genome-Wide Changes in Alternative Splicing, Inducible trans-Acting Activity of Phased Secondary Small Interfering RNAs, and Immune Responses.J Virol. 2017 May 12;91(11):e00247-17. doi: 10.1128/JVI.00247-17. Print 2017 Jun 1. J Virol. 2017. PMID: 28331096 Free PMC article.
-
Potato Spindle Tuber Viroid RNA-Templated Transcription: Factors and Regulation.Viruses. 2018 Sep 17;10(9):503. doi: 10.3390/v10090503. Viruses. 2018. PMID: 30227597 Free PMC article. Review.
-
Silencing of transcription factor encoding gene StTCP23 by small RNAs derived from the virulence modulating region of potato spindle tuber viroid is associated with symptom development in potato.PLoS Pathog. 2019 Dec 2;15(12):e1008110. doi: 10.1371/journal.ppat.1008110. eCollection 2019 Dec. PLoS Pathog. 2019. PMID: 31790500 Free PMC article.
-
Viroids: self-replicating, mobile, and fast-evolving noncoding regulatory RNAs.Wiley Interdiscip Rev RNA. 2010 Nov-Dec;1(3):362-75. doi: 10.1002/wrna.22. Wiley Interdiscip Rev RNA. 2010. PMID: 21956936 Review.
Cited by
-
In silico survey of the central conserved regions in viroids of the Pospiviroidae family for conserved asymmetric loop structures.RNA. 2019 Aug;25(8):985-1003. doi: 10.1261/rna.070409.119. Epub 2019 May 23. RNA. 2019. PMID: 31123078 Free PMC article.
-
Viroid Replication, Movement, and the Host Factors Involved.Microorganisms. 2024 Mar 12;12(3):565. doi: 10.3390/microorganisms12030565. Microorganisms. 2024. PMID: 38543616 Free PMC article. Review.
-
Symptomatic plant viroid infections in phytopathogenic fungi.Proc Natl Acad Sci U S A. 2019 Jun 25;116(26):13042-13050. doi: 10.1073/pnas.1900762116. Epub 2019 Jun 10. Proc Natl Acad Sci U S A. 2019. PMID: 31182602 Free PMC article.
-
Cellular roadmaps of viroid infection.Trends Microbiol. 2023 Nov;31(11):1179-1191. doi: 10.1016/j.tim.2023.05.014. Epub 2023 Jun 20. Trends Microbiol. 2023. PMID: 37349206 Free PMC article. Review.
-
Novel Viroid-Like RNAs Naturally Infect a Filamentous Fungus.Adv Sci (Weinh). 2023 Jan;10(3):e2204308. doi: 10.1002/advs.202204308. Epub 2022 Dec 14. Adv Sci (Weinh). 2023. PMID: 36515275 Free PMC article.
References
-
- Flores R, Minoia S, Carbonell A, Gisel A, Delgado S, Lopez-Carrasco A, Navarro B, Di Serio F. 2015. Viroids, the simplest RNA replicons: how they manipulate their hosts for being propagated and how their hosts react for containing the infection. Virus Res 209:136–145. doi:10.1016/j.virusres.2015.02.027. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
